
Бесплатный фрагмент - Предыстория человечества: от «Адама» до «Авраама»

Введение
Люди всего мира задаются вопросами: Кто мы? Кто наши предки и откуда они родом? Как долго существует человечество? По каким маршрутам передвигались наши предки до того, как оказались в современных местах обитания? Как установить происхождение отдельного человека, народа и их родство друг с другом? и т. д.
Пытаясь ответить на эти вопросы, многие люди уповают на потусторонние и сверхествественные силы, провозглашают человека венцом божественного творения, тем самым загоняя себя в тупик и признавая своё ничтожество (человек — тварь божья).
Есть категория людей, которые верят в человеческий разум, путём научного поиска стремятся познать окружающий мир, прошлое, настоящее и будущее человечества. К этой категории людей, которых можно назвать исследователями, относятся учёные, работающие в различных сферах науки. В настоящее время, науки, которые в наибольшей мере акцентированы на изучении предыстории человечества, — археология, генетика, лингвистика и др. В частности, археологи всего мира ищут признаки каменных индустрий, которые позволили бы различать среди них созданные людьми современного анатомического типа (ЛСА) (англ. Anatomically modern human, AMH, лат. Homo sapiens sapiens), неандертальцами и другими видами древних гоминидов. В последние годы достигнуты серьёзные успехи в этих поисках. Так, например, археологи видят различия в стратегиях первичного расщепления камня, применяемых нашими предками эпохи среднего палеолита, неандертальцами и денисовцами. В научной литературе всё более популярной становится идея о связи человека современного анатомического типа с индустриями, основанными на центростремительной технологии леваллуа и нубийском леваллуа.
В последние годы больших успехов достигли генетики. В частности, открытие новых методов изучения древней ДНК, позволило сравнивать геномы людей разных эпох, находить их генетических родственников в современных популяциях. Кроме того, исследование палеоДНК и современной ДНК позволяет верифицироватьь исторические реконструкции, основанные на археологических материалах.
В данной книге предпринята попытка, опираясь в основном на новейшие данные археологии и генетики заглянуть в предысторию человека современного анатомического типа от момента его формирования до появления и начала миграций представителей Y-хромосомной гаплогруппы Е. Автор не считает этот труд «истиной в последней инстанции». Где-то могут быть допущены ошибки, вызванные недостаточностью используемого материала, возможно неверным осмыслением некоторых данных, но, на мой взгляд, методология научного поиска по заданной тематике определена верно и послужит ещё многим исследователям в их благородной и трудной работе.
I. Непосредственный предшественник человека и его возраст
1.1. Возраст человека современного анатомического типа (Homo sapiens sapiens) и время его исхода из Африки
В 2000-е годы большинство специалистов считало, что Homo sapiens возник в Восточной Африке примерно 200 тыс. лет назад, а современное внеафриканское человечество происходит от группы людей, которая покинула Африку через Баб-эль-Мандебский пролив примерно 65 тыс. лет назад. С территории Южной Аравии люди стали расселяться вдоль побережья Индийского океана на восток [5, c.21].
В 2012 году, генетики, исходя из расчётов скорости мутаций у современных людей, пришли к выводу, что время исхода из Африки следует удревнить до 130—90 тыс. лет назад [37].
Результаты исследования ДНК древних людей из Южной Африки, опубликованные в 2017 году, показали, что Homo sapiens появились на Земле ранее 300 тыс. лет назад (самое раннее расхождение между человеческими популяциями могло произойти от 350 до 260 тыс. лет назад) [39]. Расчёты с помощью программного обеспечения BEAST2 показали, что время жизни последнего общего предка современных людей и неандертальцев (TMRCA) ~408 тыс. лет назад [20], т.е. мужчина, предок современного человечества, жил ~408 тыс. лет назад. Нужно отметить, что по мнению генетиков: «’«последний общий предок»», существовавший в точке генетического расхождения линий — это не конкретный индивидуум — отец или мать будущих «народов», «Адам» или «Ева», а условный момент в истории единой популяции, после которого свободный генетический обмен в ней прекращается» [6, с. 75].
Выход Homo sapiens из Африки, согласно Hershkovitz et al. (2018), мог произойти около 200 тыс. лет назад и ранее — останки человека современного анатомического типа из пещеры Мислия (Misliya Cave) в Леванте имеют возраст, полученный при совместном применении методов урановых рядов (Uranium-series, U-series) и электронно-парамагнитного резонанса (ЭПР) 174±20 тыс. лет назад (календарных) (при стандартном отклонении 2σ) [22]. Однако, Sharp, Paces (2018) утверждают, что у Hershkovitz et al. (2018) нет достоверных U-series дат старше ~70 тыс. лет назад и останки имеют минимальный возраст не старше ~60—70 тыс. лет [40]. Отметим, что у российских археологов для пещеры Мислия приведена OSL дата 130±33 тыс. лет назад (предположительно минимальная) [1, c.43].
1.2. Homo bodoensis или Homo heidelbergensis — непосредственный предок человека современного анатомического типа?
1.2.1. Гейдельбергский человек (Homo heidelbergensis)
Предком человека современного анатомического типа и неандертальцев (Homo neanderthalensis) долгое время считался гейдельбергский человек (Homo heidelbergensis) (рис. 1) (большинство исследователей придерживается этой точки зрения и поныне).

Рис. 1. Реконструкция головы взрослого Homo heidelbergensis. Выставлена в Зале происхождения человека Смитсоновского музея естественной истории в Вашингтоне, округ Колумбия (США) [23]
Название виду дано в честь г. Гейдельберг, вблизи которого (около деревни Мауэр), в земле Баден-Вюртемберг в Германии, впервые найдены останки этого гоминина. Время существования Homo neanderthalensis 800—30 тыс. лет назад. Рост Homo heidelbergensis, обнаруженных на юге Италии, судя по оставленным ими следам, не превышал 1,5 м [7].
В Европе, как полагает ряд исследователей, Homo heidelbergensis, стал предком неандертальцев, в Африке — современного человека. В Азии поздних Homo heidelbergensis (Шанидар, Табун, Тешик-Таш и др.) иногда относят к неандертальцам [2].
Wood, Collard (2000) cамым ранним представителем Homo heidelbergensis считают Бодо, который датируется примерно 600 тыс. лет назад (подробнее о Бодо, ниже). В этот вид древних людей они включают Флорисбад, Брокен Хилл (Кабве) из Замбии, Эяси из Восточной Африки, Рабат и Джебель-Ирхуд из Северной Африки. Образцы среднего возраста Homo heidelbergensis (около 400 тыс. лет назад), по их мнению, включают черепа из Хоупфилда/Эландсфонтейна (человек Салданьа) из Южной Африки, Ндуту из Танзании, Сале (Sale) из Марокко и Thomas Quarry из Марокко. Homo heidelbergensis в Азии, по их мнению, представлен свидетельствами из Нгандонга в Индонезии, Дали, Мапа и Юньсянь (Yunxian) в Китае и Хатнора (Hathnora) в Индии [42, p.142].
Васильев (2009) сближает с Homo heidelbergensis (фактически включает в этот вид) гоминида из Бодо (Эфиопия) (550—640 тыс. лет). Имеющего эректоидно-сапиентные характеристики (большой надбровный валик, толстые кости свода, высокая чешуя височной кости, значительный изгиб основания черепа, намечающаяся клыковая ямка) Бодо относили то к архаичному Homo sapiens, то к Homo heidelbergensis. Также, как и Бодо, по мнению Васильева (2009) к Homo heidelbergensis относятся гоминиды из Восточной и Южной Африки — Ндуту (370—990 тыс. лет назад), Брокен Хилл (130—300 тыс. лет), Салданья (400—700 тыс. лет), Эяси (130 тыс. лет) [4, с. 84]. К Homo heidelbergensis Васильев (2009) относит также среднеплейстоценовых поздних гоминид Африки, которых некоторые авторы выделяют в вид Homo helmei — Элие-Спрингс, Летоли 18 (около 200 тыс. лет), Омо 2, Флорисбад (260 тыс. лет), Бордер Кэйв, Мумбва и др. [4, с. 84].
В Леванте и Передней Азии Васильев (2009) относит к Homo heidelbergensis Кафзех VI и IX (115—92 тыс. лет), Схул IV и V (100—70 тыс. лет), Табун I (120 тыс. лет), Амуд I (50—40 тыс. лет), Шанидар I и II (46 тыс. лет), Кебара (61—48 тыс. лет). Обосновывает он такое мнение морфлогической мозаичностью этих людей (сочетание эректоидности с сапиентностью и/или неандерталоидностью) [4, с.85]. По мнению Васильева (2009), поскольку Передняя Азия представляет территорию, по которой постоянно проходили миграции древних людей из Африки на восток и с востока в Европу, здесь изначально Homo ergaster, а позднее Homo heidelbergensis, имея в своей морфологии некоторые сапиентные и/или неандерталоидные характеристики, формировали различные метисные варианты, максимально стабилизирующиеся только на подвидовом уровне. Исходя из этих соображений Васильев (2009) относит неандерталоидно-сапиентного Схула, эректоидно-неандерталоидных Табун, Шанидар, Амуд и эректоидно-сапиентного Кафзех VI к Homo heidelbergensis, определяя их таксономический статус по виду предшественнику, полагая, что практически во всех случаях, предшественником переднеазиатского полиморфного населения был вид Homo heidelbergensis (один из представителей, наряду с Homo erectus и Homo ergaster, эректоидных форм) [4, с. 85].
К Homo heidelbergensis могут относиться гоминиды из Китая — Дали (120—230 тыс. лет) и Цзиньнюшань (200—300 тыс. лет), а также два черепа, найденные в округе Юнсян (провинция Хубэй). Кроме того, к Homo heidelbergensis относятся люди из Нгандонга (примерно 100 тыс. лет). [4, с. 85].
Зубов (2001) описывает эволюцию гейдельбергского человека в Европе в неандертальцев и в современного человека в Африке следующим образом: «Одна из ветвей этого вида примерно 800 тыс. лет назад мигрировала в Европу и, адаптировавшись к довольно суровому климату новых мест, постепенно преобразовалась в Homo neanderthalensis, другая ветвь осталась в Африке и дала начало эволюционной линии Хоупфилд — Ндуту — Кабве — Элие Спрингс- Омо-2 — LH18 (Летоли-18 или Нгалоба; ~120 тыс. лет назад) из Танзании) и, наконец, Омо-1 (Homo sapiens). Впоследствии потомки этой линии мигрировали в Переднюю Азию, а затем — примерно 35—40 тыс. лет назад — в Европу» [8, с.].
1.2.2. Homo bodoensis
В 2022 году международная группа исследователей под руководством палеоантрополога Мирьяны Роксандич (Mirjana Roksandic) описала Homo bodoensis (рис. 2), который, по их мнению, является непосредственным предком современных людей. Вид назван по месту обнаружения черепа в 1976 г., в долине реки Бодо Д'ар в Эфиопии. Череп принадлежал человеку, жившему в эпоху среднего плейстоцена, около полумиллиона лет назад [35]. Выше показано, что этого человека также относят к самым ранним представителям вида Homo heidelbergensis [42, p.142].

Рис. 2. Художественная реконструкция Homo bodoensis (Image credit: Ettore Mazza) [16]
Homo bodoensis обитал в Северной Африке и Юго-Восточной Европе. В среднем плейстоцене, 774—129 тыс. лет назад, в Африке появился человек современного анатомического типа (Homo sapiens), а в Европе неандерталец (Homo neanderthalensis) (рис. 3) [35].
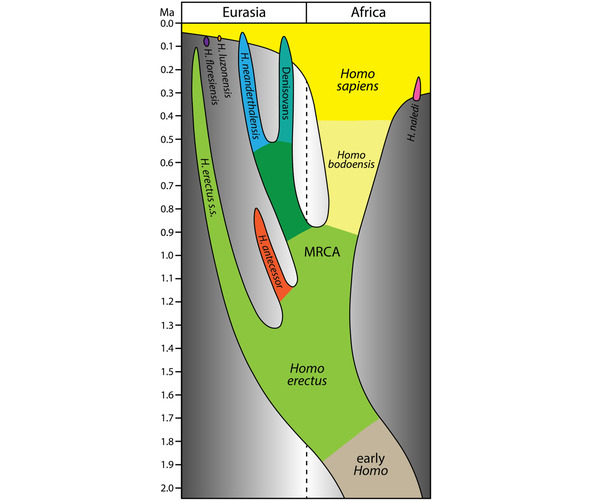
Рис. 3. Упрощенная модель эволюции рода Homo в последние 2 млн. лет, в которой Homo bodoensis позиционируется как предок (в основном в Африке) Homo Sapiens [35, Fig. 1].
По мнению Roksandic et al. (2022), к Homo bodoensis следует относить почти все находки Homo, сделанные на африканском континенте и некоторые останки, обнаруженные на юго-востоке Европы. Останки древних гоминид в Европе, классифицированные, как Homo heldelbergensis и rhodesiensis нужно считать неандертальцами [35].
Вполне возможно, что творением рук Homo bodoensis являются деревянные постройки в районе водопада Каламбо Фоллс (Kalambo Falls) в Замбии (граница Замбии и Танзании). Археологи, в заболоченных отложениях на месте археологических раскопок близ этого водопада (местонахождение Каламбо Фоллс), обнаружили два переплетающихся бревна из ивы крупноплодной (Combretum zeyheri), соединённых поперек намеренно вырезанной выемкой (обнаружены следы обработки огнём и каменными инструментами). Исследователи предполагают, что брёвна могли быть частью фундамента сооружения, дорожкой или приподнятой платформой на участке, который регулярно заливался водой.
Датирование этих брёвен методом люминесцентного датирования дало дату не менее 476±23 тыс. лет назад. Эта конструкция не имеет известных параллелей ни в африканском, ни в евразийском палеолите. Самым ранним известным деревянным артефактом является фрагмент полированной доски из ашельской стоянки Гешер Бенот Яаков, Израиль, возрастом более 780 тыс. лет назад. Деревянные орудия для собирательства и охоты появились около 400 тыс. лет назад в Европе, Китае и, возможно, в Африке. В Каламбо также обнаружили четыре деревянных орудия возрастом от 390 до 324 тыс. лет назад, в том числе клин, палку-копалку, обрезанное бревно и надрезанную ветку [13].
Человеческие останки на месте находки деревянных артефактов обнаружены не были, но исследователи предполагают, что они были созданы Homo heidelbergensis, обитавшим в этот период в данном регионе Африки [14; 17].
Исходя из возраста брёвен в Каламбо Фоллс 476 ± 23 тыс. назад, можно согласиться с тем, что их обработкой занимались Homo heidelbergensis или, если придерживаться гипотезы Roksandic et al. (2022), что на мой взгляд, более верно, Homo bodoensis, который, по их мнению, является непосредственным предком современных людей [35]. Выше упоминалось, что расчёты с помощью программного обеспечение BEAST2 показали, что время жизни последнего общего предка современных людей и неандертальцев (TMRCA) ~408 тыс. лет назад [20]. Если исходить из даты ~408 тыс. лет назад, то обитатели Каламбо Фоллс — общие предки человека современного анатомического типа и неандертальцев. В то же время, деревянные орудия, возрастом от 390 до 324 тыс. лет назад в Каламбо [13], — творение предков человека современного типа. Они созданы после ухода предков неандертальцев в Европу и, возможно, на север Африки (Джебель Ирхуд).
II. Предполагаемые Y-хромосомный «Адам» и носители Y-хромосомных гаплогрупп А00 и А0-Т и их прародина
2.1. Y-хромосомный «Адам»
По данным генетиков, всё современное человечество является потомком Y-хросомного «Адама» (ROOT (Y-Chromosome «Adam»)), предположительно жившего в Африке и возраст которого пока точно не определён (рис. 4). Нам известно, что расчёты с помощью программного обеспечение BEAST2 показали, что время жизни последнего общего предка современных людей и неандертальцев (TMRCA) ~408 тыс. лет назад [35]. Соответственно, возраст Y-хросомного «Адама» ~408 тыс. лет назад и судя по имеющимся палеонтологическим находкам он вряд ли был похож на современного человека.
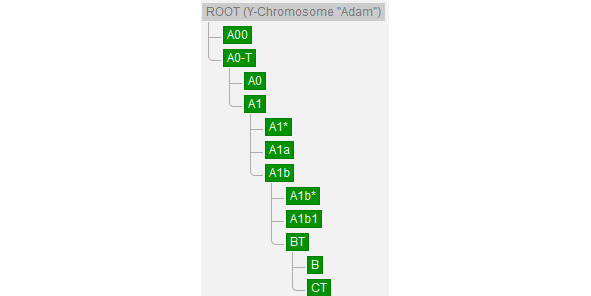
Рис. 4. «Основание» генеалогического древа современного человечества [77]
В настоящее время на Земле проживают потомки двух условных «сыновей» ROOT — A00 (гаплогруппа сформировалась 235900 (243700—228300) лет назад, время жизни последнего общего предка современных её представителей (TMRCA) 37600 (42200—33100) лет назад [78]. Представитель базальной линии этой гаплогруппы, возрастом 7885 лет назад (I10871), обнаружен на северо-западе Камеруна (археологический памятник Шум-Лака (Shum Laka)) и все известные ныне живущие представители патрилинейной наследственности A00 живут в Камеруне [78]. Подробнее о YDNA A00 изложено в параграфе, посвящённом этой гаплогруппе.
Потомками другого «сына» ROOT — A0-T (гаплогруппа сформировалась 235900 (243700—228300) лет назад, время жизни последнего общего предка современных её представителей (TMRCA) 161300 (169900—152900) лет назад), является вся остальная мужская половина современного человечества [79].
Если ориентироваться на дату ~408 тыс. лет назад (время жизни последнего общего предка современных людей и неандертальцев (TMRCA) ~408 тыс. лет назад [35]) и возраст гаплогрупп А00 и А0Т 235900 (243700—228300) лет назад [79], то можно полагать, что на протяжении 173 тыс. лет наши предки представляли единую популяцию, которая разделилась примерно 235 тыс. лет назад.
Для того, чтобы удостовериться, что изначально предки современного человечества составляли единую популяцию, можно сравнить возраст Y-хромосомных и митохондриальных гаплогрупп, представленных у современного человечества. При этом, нужно заметить, что в литературе информация о возрасте последних очень противоречива. Так, например, на сайте компании YFull на филогенетическом древе возраст митохондриальных гаплогрупп не рассчитан (рис. 5).

Рис. 5. «Основание» филогенетического древа мтДНК современного человечества (начинается с гаплогруппы L); HD — это неандертальцы. Носитель базальной линии этой гаплогруппы — неандерталец Sima de los Huesos из Испании, возрастом 394930 (149930—649930) лет назад; DN — денисовцы; NA — неандертальцы. Носитель бальной линии этой гаплогруппы неандерталец Hohlenstein-Stadel из Германии, возрастом 1237300 (61943—182490) лет назад [76]
Schlebusch et al. (2020) приводят для L возраст 179,03 тыс. лет назад, её дочерних клад L0 — 144,931 тыс. лет назад и L1’2’3’4 152,809 тыс. лет назад [61, Fig. S5.7]. У Maier et al. (2022) возраста мтДНК L — нет (у них она обозначена L0'1'2'3'4'5'6'7), но есть время жизни последнего общего предка современных представителей этой гаплогруппы — 143454 (160228—127140) лет назад [48, Table S3].
Можно ориентироваться на расчёты Ingman et al. (2000), согласно которым TMRCA для мтДНК составляет 171500±50000 лет назад [42, p.712]. Дата 171500 лет назад очень близка к времени жизни последнего общего предка современных представителей Y-хромосомной гаплогруппы A0-T (TMRCA) -161300 (169900—152900) лет назад) согласно YFull [79]. Если же мы проведём небольшой подсчёт — 171000+50000=221000 лет, то получим цифру близкую к возрасту Y-хромосомных гаплогрупп А00 и А0-Т — 235900 (243700—228300) лет назад [78; 79].
Также можно обратить внимание на время жизни последнего общего предка современных представителей Y-хромосомной гаплогруппы A0-T (TMRCA) 161300 (169900—152900) лет назад) согласно YFull [79], которое примерно совпадает со временем жизни последнего общего предка современных представителей митохондриальной гаплогруппы L0'1'2'3'4'5'6'7 по Maier et al. (2022) — 143454 (160228—127140) лет назад [48, Table S3] и возрастом клад L0 — 144,931 тыс. лет назад и L1’2’3’4 152,809 тыс. лет назад по Schlebusch et al. (2020) [61, Fig. S5.7].
Поэтому можно полагать, что наши предки, Y-хромосомный «Адам» и митохондриальная «Ева» были представителями единой популяции древних людей, обитавшей на небольшой по площади территории.
Установлено, что клады митохондриальной гаплогруппы L достигают своего пикового разнообразия в странах Восточноафриканской рифтовой долины — Эфиопии, Кении и Танзании (рис. 6) [48, Fig. 4]. Данный факт, по-видимому, является свидетельством формирования человека современного анатомического типа в этом регионе.
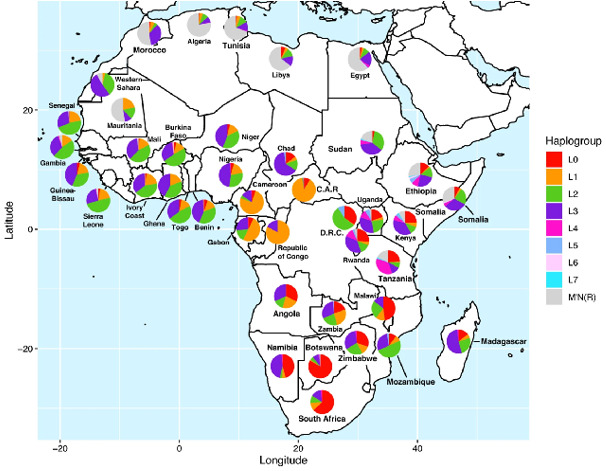
Рис. 6. Частоты распространения различных клад митохондриальной гаплогруппы L в странах Африки (L0 обозначена красным цветом) [48, Fig. 4]
2.2. Археологические культуры, представители которых могут претендовать на роль Y-хромосомного «Адама»
Выше мы пришли к выводу, что наиболее вероятной родиной человека современного анатомического типа являются страны Восточноафриканской рифтовой долины — Эфиопия, Кения и Танзания. Однако исследователи называют людьми современного анатомического типа и гоминин из других регионов, например, из Северной и Южной Африки. Поэтому кратко рассмотрим и эти версии.
2.2.1. Джебель-Ирхуд
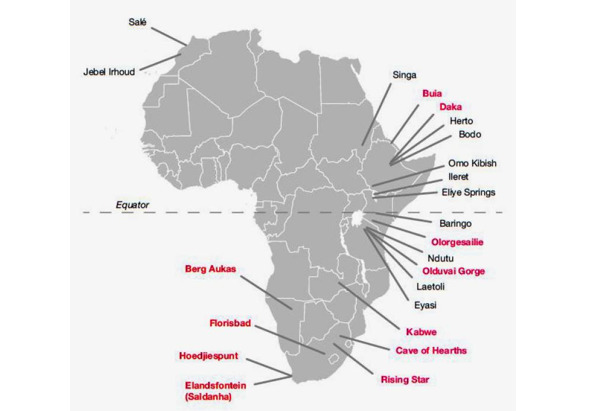
Рис. 7. Местонахождения окаменелостей ископаемых гоминид среднего и раннего плейстоцена в Африке [22]
Пещера Джебель-Ирхуд находится вблизи одноименной деревни, примерно в 100 км к западу от города Марракеш в Марокко и в 75 км от побережья (рис. 7). Она служила рудником для добычи барита (сульфата бария).
Кларк (1977) относит Джебель Ирхуд к мустье [13, с. 109]. По его мнению, мустье в Северной Африке появилось извне. Он выделяет три варианта этой традиции: один (или более) в Нубии — как с бифасами, так и без них, в Киренаике и Магрибе. «Указанные варианты отличаются разными способами техники скола: леваллуа и обработкой дисковидных нуклеусов. Доминирует техника леваллуа, поэтому эти североафриканские археологические местонахождения называются иногда „леваллуа-мустьерскими“» [13, с. 107]. Кларк (1977) пишет: «Указанные мустьерские коллекции представляют уже вполне развитую археологическую традицию, и в настоящее время в нашем распоряжении имеется мало доказательств её появления в Северной Африке в результате эволюции ашельских орудий. Это даёт возможность предполагать, что мустьерская традиция появилась в этих районах либо в результате диффузии, либо была принесена мигрантами с другого континента. Вариант из Киренаики имеет много общих черт с вариантом из Палестины, однако оба они отличаются от нубийского варианта, для которого типично большое количество зазубренных орудий, а также от магрибского варианта, который, в свою очередь, демонстрирует своеобразную региональную специфику. В это время в Северной Африке появляются некоторые евро-азиатские виды животных — медведь, европейский носорог и олень — и с ними, возможно, появляется неандертальский человек, производитель мустьерских орудий» [13, с. 109].
Hublin (2000) отмечает, что мустьерская индустрия, представленная в пещере Джебель Ирхуд, напоминает тип Ла-Феррасси (La Ferrassie) на юго-западе Франции [39, p.159], которая создана неандертальцами. В пещере Ла-Ферасси обнаружены останки трёх неандертальцев, для которых установлен возраст: погребение неандертальца Ла-Ферраси-1 (рис. 8) относится ко времени между 51,8 и 32,1 тыс. лет назад, неандерталки Ла-Ферраси-2 — 49,6—44,1 тыс. лет назад, ребёнка Ла-Ферраси-8 — 44,9—39,9 тыс. лет назад [34].
Необходимо также отметить, что Hershkovitz et al. (2018) говорят о сходстве индустрий Джебель Ирхуда и Табун D из Леванта [38].

Рис. 8. Homo neanderthalensis Ля-Ферраси 1. Мужчина 45—50 лет. Место находки: Франция, департамент Дордонь, пещера Ля-Ферраси. Датирован: 51,8 и 32,1 тыс. лет назад. Слева портрет, выполненный 3D-художником Philip Edwin [45], справа реконструкция из сети Х (пользователь InterspeciesFace) [32]
В пещере Джебель-Ирхуд обнаружены останки фауны, неатерийские археологические находки MSA (non-Aterian Middle Palaeolithic/Middle Stone Age (MSA) industries) и останки нескольких ископаемых гомининов. Ископаемые человеческие останки относятся к низшим слоям стратиграфической последовательности, наиболее известными из которых являются череп древнего человека (известен как Ирхуд 1, JI1) (рис. 9), черепная коробка взрослого индивида Ирхуд 2 (JI2) и нижняя челюсть ребёнка Ирхуд 3 (JI3) [65].
Археологические слои в Джебель Ирхуде датируются от 240±35 до 378±30 тыс. лет назад, в среднем 315±34 тыс. лет назад (термолюминесцентные даты). Непосредственно на останках ребёнка Ирхуд 3 получена дата 286±32 тыс. лет назад [41].

Рис. 9. Джебель Ирхуд 1. Портрет, выполненный 3D-дизайнером Cícero Moraes [20]
Stringer (2016) констатирует, что Ирхуд 1 и Ирхуд 2 демонстрируют некоторое фенетическое сходство с людьми из Кафзех (Qafzeh), Схула (Skhul) и Херто (Herto), хотя у них отсутствует расширение верхней теменной области. По форме свода черепа (но не лица) имеется общее сходство с окаменелостями Сима (Sima fossils) и другими ранними неандертальцами [65]. По мнению Stringer (2016), Ирхуд 1 не является анатомически современным Homo sapiens, хотя имеет намёки на «современный» базикраниальный угол изгиба в соотношении основания черепа и лица («modern’ basicranial flexion in the relationship of the face and vault) [65].
В отличие от Stringer (2016), Hublin (1992) считает обитателей Джебель Ирхуда предшественниками создателей атерийской индустрии и людьми современного анатомического типа. По его мнению, явные синапоморфии с современным человеком в сочетании с некоторыми сохранившимися плезиоморфными признаками указывают на несколько более примитивный (и более древний?) уровень, чем образцы Кафзех-Схул из Юго-Западной Азии [40].
Hublin et al. (2017) установили, что по строению лицевой части черепа ирхудцы почти не отличаются от современных людей. Однако резко отличаются от нас по форме затылка и внутреннему рельефу мозгового отдела черепа, приближаясь к Homo heidelbergensis и Homo erectus — он удлинён в переднезаднем направлении и не имеет выраженной «глобулярной» формы, характерной для современных людей. На черепе Ирхуд 1 выражен так называемый «шиньон» — вариант строения затылочной кости с резким перегибом между её верхней и нижней частями, встречающийся у архантропов и иногда у неандертальцев [41;60].
Исследователи объясняют подобные черты обитателей Джебель Ирхуда тем, что в ходе эволюции у людей сначала появились современные черты лица, а лишь затем — нынешняя форма мозга. Это предполагает, что форма мозга и его функции (когнитивные способности) развивались в рамках линии Homo sapiens [41; 60]. Они делают вывод, что мозаика особенностей, включающая морфологию лица, нижней челюсти и зубов, соотносящая людей из Джебель-Ирхуд с ранними или недавними анатомически современными людьми и более примитивная нейрокраниальная и эндокраниальная морфология, в сочетании с возрастом 315±34 тыс. лет, делают Джебель-Ирхуд старейшим местом обитания гоминидов в среднепалеолитической Африке, документирующим ранние стадии Homo sapiens. Это показывает, по их мнению, что эволюционные процессы, лежащие в основе появления человека современного анатомического типа, охватывали весь африканский континент, т.е. эти древние люди были частью обширной популяции, занимавшей всю территорию среднепалеолитической Африки [41;60].
Бахолдина (2022) не согласна с Hublin et al. (2017). Она констатирует, что «Сочетание развитого надглазничного рельефа, низкого свода и резкого перегиба затылка в комбинации с наличием клыковыми ямками позволяет диагностировать Ирхуд 1 как Homo heidelbergensis,…» [1, с. 105]. Палеонтологи Джеффри Шварц (Jeffrey Schwartz), Мария Мартинон-Торрес (María Martínon-Torres) также не согласны с тем, что останки из Джебель-Ирхуд принадлежат виду Homo sapiens, поскольку, как заметила Мартинон-Торрес, у их черепов не выражен подбородок и лоб так, как они выражены у современного человека [25].
В исследовании Mounier, Lahr (2019) Ирхуд-1 занял промежуточное положение между современными людьми и неандертальцами. Они после моделирования филогенетических связей между древними и современными черепами, выдвинули два предположения:
1) окаменелости из Марокко, по-видимому, представляют собой останки популяции, которая в среднем палеолите смешивалась с неандертальцами и не внесла вклада в появление современных людей. Основано данное предположение на том, что Ирхуд-1 показывает морфологическую близость с неандертальцами. У Ирхуда-11 нет подбородка, характерного для современного человека и имеющегося у юного Ирхуда-3;
2) альтернативная гипотеза относительно североафриканской популяции, представленной окаменелостями из Джебель-Ирхуда, может основываться на результатах геномного анализа, предполагающих, что митохондриальная ДНК (мтДНК) предков неандертальцев, общая с мтДНК окаменелостей Денисова и Сима-де-лос-Уэсос (Sima de los Huesos), была заменена африканской мтДНК между 460 и 219 тыс. лет назад. Учитывая фенотипическое сходство Ирхуд-1 как с неандертальцами, так и с ранними Homo sapiens, возможно, что окаменелости Ирхуда представляют собой местных потомков африканской популяции, которая расселилась из Африки во время событий в Зелёной Сахаре, связанных с MIS 9, и, таким образом, связаны с африканскими популяциями, которые в это время мигрировали в Европу и, следовательно, внесли свой вклад в эволюцию «классических» неандертальцев [56].
Ближе всего к истине, на мой взгляд, Mounier et al. (2019), которые считают, что Ирхуд-1 — представитель популяции Homo heidelbergensis, основная часть которой мигрировала в Европу и стала предком неандертальцев и Бахолдина (2022), согласно которой, Ирхуд 1 — это Homo heidelbergensis и соответственно, люди из пещеры Джебель-Ирхуд, относятся к этому виду древних людей.
2.2.2. Форсмит (Fauresmith)
Существует гипотеза, что прародина человека анатомически современного типа находится в Южной Африке, к югу от реки Замбези, на севере современной Ботсваны [28]. Поэтому рассмотрим индустрии MSA Южной Африки.
Форсмит (Fauresmith) — региональная южноафриканская переходная индустрия от раннего к среднему палеолиту. Индустрия/культура получила своё название по находкам близ г. Фаурсмит (Форсмит) в ЮАР. Стоянки типа Форсмит найдены также на территории Восточной Африки вплоть до Эфиопии [18].
Clark (1986) датировал форсмит в регионе Оранжевой реки между 115—80 тыс. лет до н. э. [29, p.287].
Kuman et al. (2020) пишут, что форсмит датировался 500—280 тыс. лет назад, то есть от позднего ашеля до самой ранней даты появления MSA на юге Африки, в Флорисбаде (Florisbad) — 279±47 тыс. лет назад (ЭПР-дата). Даты со стоянки Кату Пан, полученные на основании комбинации результатов OSL-датирования, U-серий и электронного спинового резонанса, показали, что индустрия форсмит древнее и появилась примерно 0,6—0,5 млн. лет назад. Возраст самых поздних комплексов индустрии форсмит в пещере Вандервер (Wonderwerk Cave) имеет сомнительную дату 187±8 тыс. лет назад, полученную на основе U-series датировки фррагмента сталагмита [44]. По мнению Kuman et al. (2020) данная индустрия развивается в результате результате изменений в поведении, связанных с развитием Homo sapiens [44].
Для индустрии форсмит характерно сочетание ашельской и леваллуазской техники: рубила, кливеры, скребки, скрёбла довольно часто встречаются, сочетаясь с леваллуазскими нуклеусами, пластинами, ретушированными остриями и орудиями геометрических форм [3].
Наиболее изученные стоянки с индустрией форсмит — Кату Пан и Кантин-Копье (Canteen Kopje), расположенные в Северо-Капской провинции ЮАР.
2.2.2.1. Кату Пан
Археологический комплекс Кату Пан представляет собой скопление нескольких археологических местонахождений, относящихся в основном к каменному веку, расположенных в окрестностях города Кату, на южной окраине пустыни Калахари, в Северо-Капской провинции ЮАР.
Кату-Пан-1
В Кату-Пан-1 (КП1) (слой 4b), обнаружена индустрия раннего палеолита. Выше него, в слое 4а, датируемом комбинацией OSL и ESR/U-серий, примерно 500 тыс. лет назад, содержится индустрия форсмит (Fauresmith), относящаяся к рубежу между ранним и средним палеолитом, которая характеризуется отщеповой технологией, изготовлением пластин и односторонних ретушированных острий, которые, по мнению археологов, представляют собой самые ранние наконечники копий. Применялись преференциальная и рекуррентная левалуазские стратегии расщепления и нелеваллуазские стратегии [72; 73; 74].
Пластинчатая индустрия форсмит местонахождения Кату Пан 1 (слой 4a), является наиболее ранней в Южной Африке. Культуросодержащий слой, в котором она обнаружена, имеет даты: ОСЛ — 464±47 тыс. лет назад. Для зуба зебры, извлечённого из этого слоя, комбинированным методом урановых серий и электрон-спин-резонанса (ESR) получены даты 542±140/107 тыс. лет назад [58].
Для вышележащего слоя 3 (относимого к среднему каменному веку) методом OSL был получен возраст 291±45 тыс. лет [72; 73; 74].
Кату-Пан-6
Местонахождение под открытым небом Кату Пан 6 (КП6) расположено примерно в 500 м к юго-востоку от КП1. В Кату Пан 6 выявлена сложная последовательность четырёх каменных индустрий, включающая ассамбляжи форсмита — слой 12 (соответствует слою 4а KП1, датируемому ~500 тыс. лет назад), раннего среднего палеолита (MSA) — слой 11 (ранний тип MSA; сопоставим с индустрией слоя 3 KП1, датируемого 291±45 тыс. лет), среднего палеолита (комплекс Ховисонс-Порт (Howiesons Poort) (HP)) — слои 9—10 и позднего палеолита ((LSA) — верхние слои; комплекс Уилтон (Wilton) с микролитическим компонентом, подобным, обнаруженному в слоях 3b–4c пещеры Вондерверк (Wonderwerk)). В Кату Пан 6 слои 7 и 8 археологически стерильны [47]. Индустрия позднего палеолита Уилтон (the Later Stone Age Wilton industry) в KП6, датируется между 5,7±0,3 тыс. лет назад и 2,3±0,1 тыс. лет назад [47].
Lukich et al. (2019) возраст раннего среднего палеолита в слоях 11 и 12 (KП6—2 и KП6—1 соответственно) определили в 120—95 тыс. лет, что значительно моложе возраста 291±45 тыс. лет для аналогичной индустрии из соседнего местонахождения KП1, датированной в 2010 году [47]. Пытаясь объяснить такую огромную разницу в датировках, Lukich et al. (2019) отмечают, что ранний каменный материал MSA как из KП1, так и из KП6 сохранился очень плохо и не был предметом детального анализа, что оставляет открытой возможность того, что это разные индустрии, которые подверглись сходным тафономическим процессам. Точно такая же ситуация с датировкой индустрии форсмит (Fauresmit) из слоя 12 KП6, ~155 тыс. лет назад, что намного моложе, чем ожидалось (возраст ассамбляжа форсмит в KП1, ~500 тыс. лет назад) [47].
2.2.2.2. Истоки индустрии MSA в Южной Африке и таксономический статус её создателей
Даты индустрии форсмит Южной Африки имеют огромный разброс дат от 542±140/107 тыс. лет назад / 464±47 тыс. лет назад (пластинчатая индустрия форсмит местонахождения Кату Пан 1 (слой 4a)) [58] до ~155 тыс. лет назад (слой 12 Кату-Пан-6, соответствующий слою 4а Кату Пан 1) [47].
Поэтому можно выдвинуть несколько предположений.
1) Наиболее вероятно, что более точны даты, полученные Lukich et al. (2019) в 2019 году [47].
Если исходить из дат Lukich et al. (2019) можно обратить внимание, что дата 155 тыс. лет назад для слоя 12 КП6 (соответствует слою 4а KП1 с индустрией форсмит, датируемому ~500 тыс. лет назад) близка к возрасту предковой для большинства современного человечества Y-хромосомной гаплогруппы А1 (сформировалась 161300 (169900—152900) лет назад, TMRCA 133400 (139000—127800) лет назад) и А0 (сформировалась 161300 (169900—152900) лет назад), встречающейся только в Африке [79]. Это может означать, что создатели индустрии форсмит были ветвью единой для современного человечества популяции, мигрировавшей с северо-запада Африки на юг этого континента и впоследствии, вымершей.
2) Если допустить, что верны даты, полученные в 2010 году Porat et al. (2010), то вполне возможно, что индустрия форсмит появилась в Южной Африке вместе с Homo erectus с северо-востока этого континента. В то же время, если правы Hershkovitz et al (2018), которые утверждают, что древнейшая индустрия Кату-Пан, имеет сходство с индустрией в Джебель-Ирхуде в Магрибе [38], гоминин из которого, скорее всего, Homo heidelbergensis [1, с. 105], то и создатели южноафриканской индустрии в Кату-Пан, должны относиться к этому виду древних людей.
Подтверждением того, что создатели индустрий MSA пришли в Южную Африку с севера, служит тот факт, что в Восточной Африке переход от ашеля к MSA произошёл до 284 тыс. лет назад в формации Каптурин в Кении. В той же Кении, в Олоргесайли (Olorgesailie), комплексы, датируемые около 615—499 тыс. лет назад, имеют черты, отличающие их от более старых ашельских находок на том же участке и более характерные для MSA, такие как небольшие нуклеусы (small cores) (с некоторых из них были сняты несколько пластинок), мелкие отщепы и более экзотические источники камня, или акцент на одном источнике камня более высокого качества. Переход к MSA, в котором отсутствуют ашельские элементы, датируется на этом местонахождении 305—320 тыс. лет назад [44].
Древнейшая дата для MSA Южной Африки — 279±47 тыс. лет назад происходит с местонахождения Флорисбад [3]. Гоминид Флорисбад (Florisbad), живший здесь 259±35 тыс. лет назад [56, Table 1], отнесён к виду Homo heidelbergensis [2, с. 84].
В пещере Бордер (Border Cave) на границе Свазилэнда и ЮАР (провинция Квазулу-Наталь) ЭПР-даты для древнейших слоёв MSA указывают на возраст свыше 200 тыс. лет [3]. В пещере 13В в Пиннакл-Пойнт на южном побережье Мыса Доброй Надежды древнейшие слои MSA имеют даты порядка 180 тыс. лет [3], ~164±12 тыс. лет [49]. Здесь найдены окаменелости двух гомининов. Обнаруженный резец метрически занимает промежуточное положение между гомининами среднего плейстоцена и современными людьми. Каменные артефакты типологически и технологически напоминают региональный вариант MSA (индустрия залива Моссель), но значительно меньше [50].
В пещере Вандерверк возраст индустриии MSA, согласно результатам ТУ-датирования, составляет 150–200 тыс. лет [3]. Останки гоминин не известны.
Кроме того, южноафриканская стоянка Дюйнефонтейн 2 (Duinefontein 2), относящаяся к позднему ашельскому периоду, может быть переходной. ОСЛ-даты для песков, подстилающих и перекрывающих верхний ашельский слой этого памятника, говорят о том, что он не древнее 290 и не моложе 270 тыс. лет назад, а фауна соответствует возрастному диапазону 250—300 тыс. лет назад. Однако диагностические ашельские типы орудий опубликованные с этой стоянки, ограничены одним целым и двумя сломанными рубилами, а также несколькими фрагментами рубила, и поэтому ассамбляж может быть переходным [44].
2.2.3. Олоргесайли (Olorgesailie)
Открытое местонахождение Олоргесайли расположено между двумя потухшими вулканами примерно в 70 км от Найроби на юге Кении, в восточной части Рифтовой долины (рис. 7). Названо в честь горы Олоргесайли, расположенной неподалёку. Здесь, вместе с ашельскими рубилами, обнаружены фрагменты свода черепа Homo erectus, названного KNM-OL 45500, датируемые 900—970 тыс. лет (40Ar/39Ar, магнитостратиграфия, стратиграфия) [33]. Отмечается близость KNM-OL 45500 по размерам к образцам из Дманиси (Грузия) и Homo naledi [55]. Внешность Homo erectus представлена на рис. 10).
Первые свидетельства существования человека на этой территории датируются 1,2 млн. лет назад. Исследование показало, что обитатели Олоргесайли изготавливали на протяжении 700 тыс. лет из местного сырья рубила. 305—320 тыс. лет назад у них произошёл резкий сдвиг в поведении и стратегиях выживания, появились новые технологии изготовления орудий, признаки обмена с другими группами людей и использование в быту цветных пигментов. В частности, орудия, в отличие от универсальных рубил, стали более специализированными. Многие из них представляла собой наконечники, предназначенные для прикрепления к древку и, по-видимому, использовавшиеся в качестве метательного оружия. Часть орудий имела форму скребков или шил. Небольшие каменные наконечники были изготовлены из обсидиана, источники которых находились на расстоянии 24—88 км (15—55 миль) от Олоргесайли, где нет собственного источника этого камня. Это указывает на то, что между различными группами людей в Африке мог существовать обмен. Были также обнаружены чёрные и красные камни — марганец и охра, а также свидетельства того, что камни были обработаны для использования в качестве красящего материала.

Рис. 10. Скульптура женщины Homo erectus, выставленная в Зале происхождения человека Смитсоновского национального музея естественной истории (Вашингтон, округ Колумбия, США) (реконструкция J.Gurche; фотограф T. Evanson) [57]
Проанализировав образцы горной породы, исследователи установили причины этих поведенческих, технологических и культурных сдвигов у обитателей Олоргесайли. Геологические, геохимические, палеоботанические и фаунистические данные показали, что около 360 тыс. лет назад в регионе начался длительный период нестабильности климата, землетрясения постоянно меняли ландшафт и соответвенно, менялись флора и фауна. Всё это привело к слому привычного образа жизни древних людей. Поэтому они были вынуждены приспосабливаться к непредсказуемым условиям жизни. Технологические инновации, сети социального обмена и ранняя символическая коммуникация стали теми факторами, которые помогли древним людям выжить и получить необходимые им ресурсы [24; 30; 59].
2.2.4. Каптурин
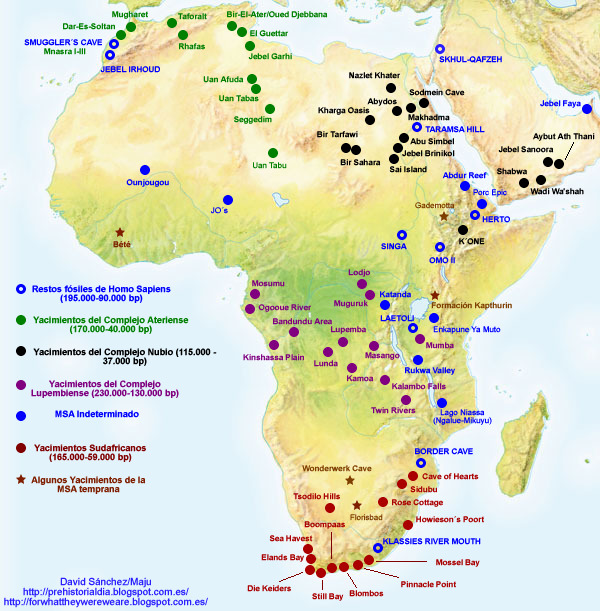
Рис. 11. Местоположение формации Каптурин и других местонахождений MSA [21]
Формация Каптурин (Kapthurin Formation) находится на холмах Туген (Tugen Hills) в Кенийской рифтовой долине (Kenya Rift Valley) (рис. 11) и представляет собой серию осадочных и вулканических отложений среднего и позднего плейстоцена толщиной 125 м, обнажившихся на площади более 150 км² к западу от озера Баринго. Формация названа в честь врезающейся в неё реки Каптурин. Слои тефры были датированы, пронумерованы снизу доверху от K1 до K5 (рис. 12) и коррелированы, чтобы обеспечить надёжную хронологию горизонтов Каптурина. В ней сохранилось множество археологических и палеонтологических памятников [31; 43].
В формации Каптурин десять археологических памятников, среди которых GnJh-41 GnJh-42 и GnJh-57, находятся в слоях, датированных радиометрическим методом (⁴⁰Ar/³ꝰAr) ~0,55–0,52 млн. лет назад (545±3 и 509±9 тыс. лет назад) (рис. 12). Их ассамбляж представлен отщепами, изготовленными из небольших галек (informal flake industry made on small cobbles). Ручные рубила (handaxes) редки, составляя лишь 1,1% или даже 0,5% от общего числа артефактов. Редкость рубил примечательна, поскольку в этот период ашельская индустрия, характеризующаяся рубилами, была широко распространена в Восточной Африке [31].
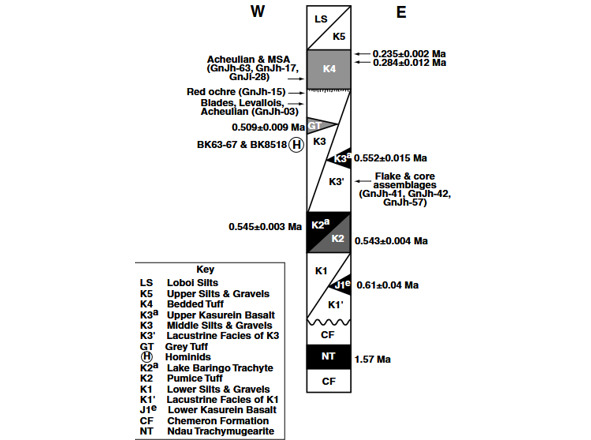
Рис 12. Схематический стратиграфический разрез формации Каптурин [31, Fig. 2].
На стоянках GnJh-42 и GnJh-50 формации Каптурин в культуросодержащих слоях, датированных (⁴⁰Ar/³ꝰAr) 545±3 и 509±9 тыс. лет назад, зафиксировано наиболее ранее в мире появление пластинчатой индустрии [43]. Отметим, что пластины в формации Каптурин появились значительно раньше, чем произошло расхождение Homo neanderthalensis и Homo sapiens [43] (расчётное время жизни последнего общего предка современных людей и неандертальцев (TMRCA) ~408 тыс. лет назад [35]). Оформление нуклеусов и скалывание пластин на этих стоянках типологически близко, по мнению исследователей, к объёмному методу, применявшемуся на стоянке Хуммала в Центральной Сирии [43, p.195].
Johnson, McBrearty (2010) отмечают, что стандартизированные заготовки, обнаруженные на стоянках GnJh-42 и GnJh-50 формации Каптурин, имеют параллельные или почти параллельные стороны и соотношение длины к ширине, равное или превышающее 2:1. Подобные стандартизированные заготовки встречаются в разном количестве на многих европейских поселениях эпохи верхнего палеолита (UP; 40 тыс. лет назад) и африканского позднего каменного века (LSA; 50 тыс. лет назад) [43, p.193]. Если это действительно так, то в обитателях стоянок GnJh-42 и GnJh-50 формации Каптурин, можно видеть древнейших прямых предков современного человечества.
2.2.4.1. Гоминиды формации Каптурин
В формации Каптурин обнаружены останки гоминид KNM-BK 63—67 и KNM-BK 8518 в слоях с датами ~543±9 и 509±4 тыс. лет назад, соответственно. Учитывая их стратиграфическое положение на 0,7 и 3 м ниже пемзового туфа (Pumice Tuff), их возраст составляет 510–512 тыс. лет назад. Эти гоминиды среднего плейстоцена являются ближайшими современниками гоминидов Бодо (датирован методом ⁴⁰Ar/³ꝰAr в интервале 640–550 тыс. лет назад) из Эфиопии, и ОН 11, ОН 23 и Ндуту из ущелья Олдувай в Танзании, возраст, которых оценивается в 490–780 тыс. лет назад [31].
Нижние челюсти KNM-BK-67 и KNM-BK-8518 имеют в целом архаичную морфологию и у них отсутствуют подбородки, что исключает их из Homo sapiens. Правая плюсневая кость KNM-BK-63 и проксимальная фаланга KNM-BK-64 также имеют архаичный внешний вид. KNM-BK-65 демонстрирует как архаичные особенности, так и признаки, которые можно увидеть у Homo sapiens. Таксономически их определяют, как Homo erectus, «архаичный» Homo sapiens, Homo rhodesiensis [31].
Как уже упоминалось, расчёты с помощью программного обеспечение BEAST2 показали, что время жизни последнего общего предка современных людей и неандертальцев (TMRCA) ~408 тыс. лет назад [35]. Гоминиды формации Каптурин, если их возраст определён верно, старше Y-хромосомного «Адама» примерно на 100 тыс. лет. Вполне возможно, что популяция, обитавшая в формации Каптурин в этот период — предковая для людей современного анатомического типа и неандартальцев (и Nesher Ramla Homo из Леванта, датируемого 140–120 тыс. лет назад (пещера Нешер Рамла) и др.). Вероятно, о развитии этой популяции в сторону сапиентации является наличие у KNM-BK-65 как архаичных особенностей, так и признаков, которые можно увидеть у Homo sapiens [31].
На принадлежность популяции древних людей из формации Каптурин к общим предкам современного человека и неандертальцев, а также Nesher Ramla Homo, косвенно указывают археологические данные. Так, например, по мнению Деревянко (2016) первичное пластинчатое расщепление, техника леваллуа (протолеваллуа), из Восточной Африки проникли в Левант, где известны на стоянке Гешер Бенот Яаков и более ранних местонахождениях [5]. Однако комплексы Каптурин «и стоянки Гешер Бенот Яаков имеют мало общего, их невозможно объединить в один комплекс» [5]. Объясняет Деревянко (2016) это следующим образом: «любые технологические традиции во время длительных передвижений изменяются в связи с необходимостью адаптации мигрирующих популяций к новым экологическим условиям (другой климат, животный и растительный мир, исходный материал для изготовления каменных орудий и т.д.), а также под влиянием технологических традиций автохтонного населения» [5, с.16].
Пластинчатая технологии, впервые зафиксированная в Каптурин, получила широкое распространение в Леванте на ашело-ябрудийском этапе позднего ашеля. В ярко выраженной форме пластинчатое расщепление прослеживается в амудийской индустрии на местонахождении Кесем в Израиле (400–200 тыс. лет назад) [5, с. 22].
2.2.4.2. Эволюционное развитие индустрий формации Каптурин в ашельский период. Каптурин — родина технологии центростремительного леваллуа, пластинчатого производства и символической деятельности
Средняя часть формации Каптурин датируется от 0,509±0,009 до 0,284±0,012 млн. лет назад. В отложениях этого возраста (стоянки GnJh-17, GnJh-63 и GnJi-28 [67]) найдены острия/наконечники (points), типичные для среднего каменного века (MSA). Эти даты старше возраста MSA в других частях Восточной Африки на 49 тыс. лет (датируется ~234 тыс. лет назад) и устанавливают возраст перехода от ашеля к MSA в регионе [31]. В этом же интервале встречаются свидетельства использования леваллуазской техники изготовления как мелких отщепов, так и заготовок бифасов, систематического изготовления пластинок (blades), использования и обработки красной охры. Присутствие пластин (blades) и красной охры важно, поскольку пластины означают высокую степень технической компетентности, а красная охра предполагает символическое поведение [31].
Красная охра обнаружена на стоянке GnJh-15, расположенной на береговом склоне р. Бартекечо, притока р. Каптурин и лежит непосредственно под слоем туфа К4, датируемого 0,284±0,012 млн. лет назад. Красная охра с этой стоянки значительно старше охры со среднепалеолитических памятников Южной Африки, таких как пещеры на реке Класье (Klasies River Caves), датированных различными методами до 120 тыс. лет назад, пещера Бломбос, имеющей дату — до 70 тыс. лет назад и Твин-Риверс (Twin Rivers) в Замбии, датированной до ~230 тыс. лет назад [31].
Сравнение больших леваллуазских отщепов, полученных в результате центростремительного расщепления галек из лавы и выработанных из них путём ретуширования ручных рубил (handaxes), кливеров (cleavers) и других подобных орудий со стоянки LHA/GnJh-03 возрастом около 400 тыс. лет назад (395.6±3.5 тыс. лет назад) с находками из четырёх классических ашельских комплексов центральной Рифтовой долины: Кариандуси (Kariandusi), Олоргесайли CL1—1 (Olorgesailie CL1—1), Олоргесайли Пачка 6/7 (Olorgesailie Member 6/7), и Исинья (Isinya) (рис. 13), выполненных на крупных отщепах, показало, что орудия формации Каптурин отличаются своей тонкостью (как и в формации Каптурин, все рубила и кливера на этих стоянках были изготовлены из мелкозернистой вулканической породы). Это достигалось за счёт симметричной центростремительной обработки пологих выпуклостей верхней поверхности нуклеусов Леваллуа, созданием выступающей гранённой платформы и проксимального скоса верхней поверхности. Кроме того, ашельские рубила и кливера формации Каптурин сравнивались с позднеашельскими подобными орудиями из Каламбо-Фоллс (Kalambo Falls), изготовленными из кварцита. Ни в одном из сравнимаемых ассамбляжей нет свидетельств леваллуазской технологии создания рубил и кливеров [62].
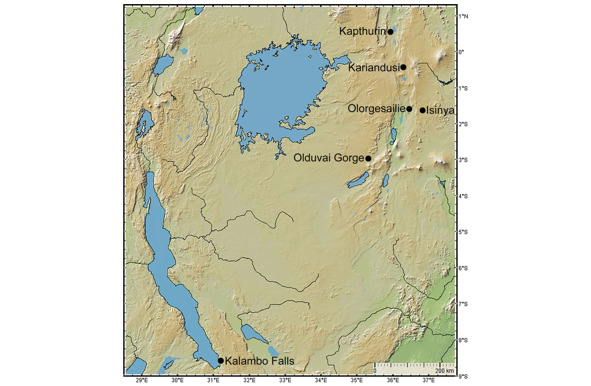
Рис. 13. Местоположение Олдувайского ущелья и ашельских комплексов, сравнимаемых с ашельским комплексом формации Каптурин [62, Fig. 1].
Леваллуазская технология из расположенной выше LHA/GnJh-03 стоянки Коимилот (Koimilot (GnJh-74); возраст на основании геохимической корреляции с датированной вулканической тефрой, составляет около 200 тыс. лет [66]; 200—250 тыс. лет [67]) скорее показывает различия в проявлении технологии производства леваллуазских отщепов, чем в типологии, что позволяет предположить развитие местной предшествующей ашельской традиции [66; 67], т.е. традиции леваллуа, зародившейся на ашельской стоянке LHA/GnJh-03, возрастом ~400 тыс. лет назад.
Первоначально McBrearty с соавторами (McBrearty et al. (1996), McBrearty, Brooks (2000)) считали, что позднеашельская пластинчатая индустрия на стоянках GnJh 3, 15, 17 формации Каптурин периода около 280 тыс. лет назад, по технологическим показателям не имела преемственной связи с более древней, возрастом свыше 500 тыс. лет [51; 52]. Позже она с соавтором (Tryon, McBrearty (2006) предположила, что различные методы производства леваллуазских отщепов в пластах, датируемых ~200–250 тыс. лет назад на стоянке Коимилот (Koimilot (GnJh-74), могли возникнуть на основе предшествующей ашельской технологии [67]. Ещё позже она с соавтором (Johnson, McBrearty (2010)) пришла к выводу, что пластины из местонахождений GnJh-42 и GnJh-50, датируемых 545—509 тыс. лет назад и GnJh-03, GnJh-17, GnJh-15, датируемых 509—285 тыс. лет назад, принадлежат к одной традиции, которая сохранялась в Восточной Африке на протяжении большей части среднего плейстоцена [43, p. 198]. Johnson, McBrearty (2010) отмечают, что географически распространённый ашельский технокомплекс, как однородная технологическая традиция, оставался устойчивым к изменениям на протяжении всего своего длительного существования, вплоть до замены индустриями MSA. Эти изменения, по-видимому, связаны с расширением поведенческого репертуара гоминин, включившего в себя действия, необходимые для производства пластин, отщепов и наконечников Леваллуа, а также бифасиально отретушированных наконечников, характерных для MSA. Эти технологические инновации, возможно, в какой-то степени повлияли на анатомические изменения, обнаруженные в палеонтологической летописи гомининов [43, p. 198].
2.2.4.3. Сосуществование популяций древних людей разных видов в формации Каптурин
Tryon, McBrearty (2002, 2006) констатируют, что в формации Каптурин хронологический порядок расположения памятников не показывает чёткого однонаправленного перехода от ашельского периода к MSA. Например, на стоянках GnJh-17 и GnJh-63 (датируются 284±12 — 235±2 тыс. лет назад) видна взаимная стратификация характерных для MSA острий/наконечников (points) и ашельских рубил (handaxes). Наконечники (points) и рубила (handaxes) также встречаются на одном и том же археологическом уровне в GnJh-17 и Ророп-Лингопе (Rorop Lingop, GnJi-28) (датируются 284±12 — 235±2 тыс. лет назад). На местонахождении 108 (Loc. 108) обнаружены кливера (cleavers) в слоях, залегающих над верхними базальтовыми туфами, и в совокупности эти наблюдения свидетельствуют о том, что ашельское производство орудий труда сохранялось и после появления наконечников. Исследователи также отмечают, что, возможно, в формации Каптурин ашельские элементы переслаиваются с сангоанскими, MSA и, возможно, с компонентами форсмита (Fauresmith). В отличие от таких местонахождений как Каламбо-Фоллс (Kalambo Falls), Пещера очагов (Cave of Hearths), Исимила (Isimila) или Мугурук (Muguruk), слои формации Каптурин, содержащие ашельские артефакты (рубила (handaxes)) лежат поверх тех, которые содержат артефакты, определяющие Сангоан (кирки (picks)) и MSA (острия/наконечники (points)). Кирки, являющиеся отличительной чертой сангоана, также встречаются в бесспорно ашельских коллекциях и ассамбляжах MSA. Так, например, в местонахождении GnJh-17 на одном уровне обнаружены рубило, кирка и наконечник. Аналогичные находки известны на уровне 7 Гарба III в Мелка-Контуре [67; 68].
По мнению Tryon, McBrearty (2002), остаётся неясным, является ли технологическое разнообразие, наблюдаемое в пределах формации Каптурин, результатом сложной адаптации гоминид к разнообразной палеосреде в пределах одного бассейна или результатом присутствия здесь нескольких видов гоминид [67]. Они предлагают несколько интерпретаций данному явлению. Во-первых, сочетание артефактов на таких стоянках, как GnJh-17, может представлять собой вариации в позднем ашеле или раннем MSA, отражающие адаптацию к среде обитания или видам деятельности. Во-вторых, они могут свидетельствовать о сохранении более древних методов обработки камня в процессе накопления новых технологий в рамках одной традиции. В-третьих, различные классы артефактов могут обозначать отдельные занятия различных групп современных гоминидов, использующих разные технологии. В-четвёртых, они могут представлять собой палимпсест отложений, накопленных с течением времени [67].
Выводы.
1) Датировки появления леваллуазской техники, использования и обработки красной охры (стоянка GnJh-15), острий/наконечников (points), типичных для среднего каменного века (MSA) (стоянки GnJh-17, GnJh-63 и GnJi-28) в Каптурине ~284 тыс. лет назад более ранние, чем появление подобных артефактов в других частях Восточной Африки ~234 тыс. лет назад. При этом, дата ~234 тыс. лет назад близка к возрасту гаплогрупп А00 и А0-Т — 235900 (243700—228300) лет назад [79]. Это позволяет предположить, что в формации Каптурин обитала предковая для всего современного человечества популяция древних людей.
2) На возможность обитания в формации Каптурин предковой для современного человека популяция Homo heidelbergensis/Homo bodoensis 545±3 — 509±9 тыс. лет назад указывает тот факт, что эти древние люди отличались от остальных популяций ашельского периода Африки орудийным набором (редкость рубил [31]). Здесь же, на стоянках GnJh-42 и GnJh-50, датируемых 545±3 — 509±9 тыс. лет назад, зафиксировано наиболее ранее в мире появление пластинчатой индустрии [43]. Впоследствии стандартизированные заготовки, подобные обнаруженным на стоянках GnJh-42 и GnJh-50 формации Каптурин встречаются в разном количестве на многих европейских поселениях эпохи верхнего палеолита (UP; 40 тыс. лет назад) и африканского позднего каменного века (LSA; 50 тыс. лет назад) [43, p.193].
3) Технология изготовления каменных орудий на ряде стоянок формации Каптурин, датируемых 509—285 тыс. лет назад, ~200–250 тыс. лет принадлежит к традиции, датируемой 545—509 тыс. лет назад [43, p. 198; 68], что может быть свидетельством эволюционного развития в данной формации Homo heidelbergensis/Homo bodoensis в Homo sapiens sapiens.
4) Примерно 200 тыс. лет назад в формации Каптурин появилось новое население. Об этом свидетельствует то, что здесь в период MSA, как полагают археологи, можно выделить два среднепалеолитических локуса, датируемые ∼200 тыс. лет назад. Для индустрии локуса 1 характерны отщепы, преференциальная и рекуррентная леваллуазские стратегии, а также стратегии, не связанные с леваллуа. Для индустрии локуса 2 характерны острия и пластины (blades), снятие пластин с нуклеусов, острия и однонаправленная конвергентная леваллуазская стратегия [66; 68; 69]. Археологи также отмечают появление на одном археологическом уровне орудий характерных для ашеля, сангоана, MSA и, возможно, форсмита (Fauresmith) [67; 68].
Вполне возможно, что в этот период в формации Каптурин началось смешение гоминидов разных видов. Есть гипотеза Mounier, Mirazón Lahr (2019), что люди современного типа, расселившиеся по планете и живущие на ней сейчас, получились в результате смешивания популяций их предшественников из Южной, и, возможно, Восточной Африки [56] (см. ниже: Гипотеза Mounier, Lahr (2019)).
5) Приход людей другого вида, по всей вероятности, привёл к их смешению с аборигенами формации Каптурин (см. ниже), а другая часть аборигенов, предположительно, могла мигрировать на север, в Гадемотта/Кулькулетти. Таким образом, из мигрантов в Гадемотта/Кулькулетти могла сформироваться популяция носителей Y-хромосомной гаплогруппы А0-Т, а в Каптурине — популяция носителей Y-хромосомной гаплогруппы А00. Впоследствии представители последней, под давлением потомков А0-Т мигрировали по зоне экваториальных лесов на запад Африки, где до сих пор есть носители YДНК А00.
2.2.5. Гипотеза Mounier, Lahr (2019) [56]
Mounier, Mirazón Lahr (2019) cмоделировали филогенетические связи между древними и современными черепами, создав трёхмерную реконструкцию древних останков и проанализировав морфологию черепов разных видов Homo. Среди них были 263 черепа, принадлежавших современным жителям планеты из 21 популяции; останки древнейших людей, человека умелого (Homo habilis), человека работающего (Homo ergaster) и дманисского гоминида, живших 1,8 и 1,6 миллиона лет назад на территории современной Кении и 1,77 миллиона лет назад на территории Грузии; останки неандертальцев, живших на Ближнем Востоке, в Западной и Южной Европе, и древнейших людей современного типа, а также черепа из Омо Кибиш, Марокко и ЮАР. Моделирование показало, что, по-видимому, 200—350 тысяч лет назад образовались несколько локальных популяций предшественников современных людей. Они мигрировали внутри континента и смешивались между собой. В результате, 100—200 тысяч лет назад, появились люди современного типа. При этом вклад различных древних популяций был неодинаковым, часть из них исчезла, не оставив следов ни в геноме, ни в фенотипе современных людей.
Основываясь на известных останках, авторы делают вывод, что люди современного типа, расселившиеся по планете и живущие на ней сейчас, получились в результате смешивания популяций их предшественников из Южной, и, возможно, Восточной Африки [56].
В исследовании Mounier, Lahr (2019) южноафриканский Флорисбад и, в меньшей степени, восточноафриканские KNM-ES 11693 и Omo II, показали наибольшую близость к современным людям. Они классифицируют череп Флорисбада как пример раннего Homo sapiens, который, как они предполагают, возник между 350 и 260 тыс. лет назад в результате смешения популяций из Восточной и Южной Африки [56].
Если судить по возрасту этих древних людей, Омо II — 195±95 тыс. лет назад; KNM-ES 11693 — 270–300 тыс. лет назад; Florisbad — 259±35 тыс. лет назад [56, Table 1], то эти древние люди пришли из Южной Африки. Однако, Omo I из формации Кибиш, в Эфиопии (Kibish formation, Ethiopia), считающийся древнейшим человеком современного анатомического типа, имеет возраст — 233±22 тыс. лет назад [70]. К тому же и сами Mounier, Lahr (2019) отмечают, что Омо I из формации Кибиш в Эфиопии (возраст: 233±22 тыс. лет назад) и Херто из формации Бури в Эфиопии (возраст: 154±7 — 160±2 тыс. лет назад) имеют анатомию, подобную современной. Все остальные африканские окаменелости LMP демонстрируют мозаику совренных и архаичных признаков — это Джебель Ирхуд, Гомде, Нгалоба, Элие Спрингс, Флорисбад, Omo II, возможно, Синга [56].
Отметим, что отсутствие в модели Mounier, Lahr (2019) черепов Омо 1 и Херто, которые, по мнению большинства исследователей, принадлежат к древнейшим представителям человека современного анатомического типа, могло очень сильно повлиять на результаты их исследования.
2.2.6. Существовал ли Homo helmei и есть ли связь между ним и (прото) /австралоидным комплексом?
McBrearty, Brooks (2000) [52] полагают, что происхождение нашего вида связано с появлением технологии среднего каменного века (Middle Stone Age, MSA) 250–300 тыс. лет назад. При этом, появление технологий среднего каменного века и первые признаки современного поведения совпадают с появлением окаменелостей, приписываемых Homo helmei, являющимся промежуточным звеном (видом) между Homo erectus/ergaster и Homo sapiens [52].
К виду Homo helmei McBrearty, Brooks (2000) относят: Флорисбад (~259 тыс. лет назад) из Южной Африки; LH18 (Летоли 18 или Нгалоба; ~120 тыс. лет назад) из Танзании; Илерет KNM-ER 3884 (Гуомде) (~270—300 тыс. лет назад) из Кении; Омо-2 (~130 тыс. лет) из Омо Кибиш в Эфиопии; Синга (133 тыс. лет назад) из Судана; Джебель Ирхуд (~200 тыс. лет; в настоящее время датируется ~315 ± 34 тыс. лет назад) из Марокко; Хауа Фтеах; Дире-Дауа (Porc-Epic); Элие Спрингс. Краниологический тип Homo helmei они определяют, как массивный, долихокранный с сильно выраженным надбровьем, уплощенной чешуей лобной кости, низколиций, низкоорбитный, прогнатный [52].
Судя по вышеприведённым датировкам можно предположить, что движение Homo helmei началось в Южной Африке (Флорисбад; ~259 тыс. лет назад) и достигло Cеверо-Восточной Африки ~130 тыс. лет (Омо 2). Однако не всё так однозначно. Так, например, согласно результатам последних исследований Ирхуд 1 — Homo heidelbergensis [1, с.105], а археологические слои в Джебель Ирхуде датируются от 240±35 до 378±30 тыс. лет назад, в среднем 315±34 тыс. лет назад (термолюминесцентные даты). Непосредственно на останках ребёнка Ирхуд 3 получена дата 286±32 тыс. лет назад [41; 60], т.е. уже исходя из датировок Джебель Ирхуда нельзя предполагать движение предков современного человека из Южной Африки на север это материка.
Омо II, для которого у McBrearty, Brooks (2000) указан возраст ~130 тыс. лет, в настоящее время датируется 195±95 тыс. лет назад [56, Table 1].
Люди из Хауа Фтех имеют возраст ~70 тыс. лет назад [65] и по своим морфологическим особенностям наиболее тесно связаны с табунской группой неандертальцев с горы Кармель [53]. Отметим, что в настоящее время упомянутые выше представители Табун С, предположительно относятся к отдельному виду людей Homo Nesher Ramla, сформировавшемуся на Ближнем Востоке и который мог заселять Юго-Западную Азию в промежутке между 420 и 120 тыс. лет назад [37].
Эти данные не свидетельствуют в пользу существования вида Homo helmei.
А.А.Зубов полагает, что в сочетании признаков, характеризующих тип Флорисбад и вид Homo helmei, чётко проступает австралоидный комплекс и Homo helmei был панэйкуменным субстратным пластом на поздних этапах формирования полиморфного вида Homo sapiens. По его мнению, черты Homo helmei обнаруживают преемственность в морфологии ископаемых черепов Австралии (например, экземпляры из Вилландра Лейкс (WLH 50), Кау Свэмп, Талгай, Кохуна) и австралоидных по типу черепов в Китае (Маба, Дали, Цзиннюшан), Вьетнаме (Май-да-Нуок), в Индонезии (Нгандонг, Вадьяк), в Индии (Нармада), на Ближнем Востоке (Зуттие), отражающих субстратный тип «восточного» надрасового «ствола». Кроме того, позднеплейстоценовые-раннеголоценовые черепа Южной Америки («палеоамериканский комплекс»), например, позднеплейстоценовый череп из Отовало (Эквадор), также несут черты описываемого типа. Поэтому он считает, что «морфология Homo helmei внесла свой вклад в формирование преимущественно протоавстралоидной ветви дифференциации ранних сапиенсов» [9]. А.А.Зубов также полагает, что Homo helmei стоял у истоков эволюции современного человека. На это указывают, по его мнению, низкое лицо, низкие орбиты и долихокрания кроманьонцев; сильно выраженный рельеф надбровья в европейской верхнепалеолитической черепной серии Младеч. А.А.Зубов считает, что «в Юго-Восточной и Восточной Азии по многим краниологическим данным этот комплекс предшествовал более позднему — монголоидному и был древнейшим субстратом для всех ветвей последнего, включая северных монголоидов и североамериканских индейцев. Австралоидный тип, […], послужил основой формирования типов аборигенного населения Южной Америки. В Африке, […] черты его можно проследить среди некоторых современных групп, в частности — в краниологическом типе готтентотов» [9].
Дробышевский (2021) полностью опровергает построения А.А.Зубова. Рассматривая проблему австралоидности, Он полагает, что из-за возраста в 259 тыс. лет назад, Флорисбад из Южной Африки, некорретно сравнивать с современными расами. Кроме того, «и морфологически описание Флорисбада как австралоида крайне спорно: его надбровный валик равномерно утолщён по всей своей протяжённости, а лицевой скелет реконструирован в большей степени, чем сохранился. Вопреки распространённой реконструкции верхняя челюсть, скорее всего, имела значительную высоту, в противоположность австралийцам. По тем же причинам в настоящее время забавным выглядит определение как „прото-австралоидов“ фрагментов из Эяси и Дире Дава» [8]. Также некорретно сравнивать с современными расами Дали из Китая, имеющего возраст около 270 тыс. лет назад [8].
Дробышевский (2021) полагает, что австралоидность достоверно отмечена на множестве верхнепалеолитических и голоценовых черепов из Индокитая и Индонезии, например, из местонахождений Конмонг, Ханг Гон — Дау Гиай, Ханг Муои, Ханг Чо, Куиньван, Май-да-Диеу, Май-да-Нуок, Тампонг, Да-Бут, Соли, Тхиеу-Зыонг, Мох Хиеу, Вадьяк, Гуа Лава, Ниа, Лянг Тоге, Табон. «Австралоидность» мезолитических людей из Белланбанди Паласса на Шри-Ланке является в действительности веддоидностью в массивном исполнении. Видимо, так же стоит оценивать «австралоидность» фрагментов из Гар-и-Камарбанд в Иране и Хараппы в Пакистане. В Африке серий черепов с «австралоидным комплексом» не найдено [8].
III. Гибридизация человека современного анатомического типа с другими видами древних людей в Африке и археологические культуры/индустрии, создатели которых могли скрещиваться с человеком современного анатомического типа
3.1. Гибридизация человека современного анатомического типа с другими видами древних людей в Африке
Hammer et al. (2011) исследовав сан (койсанский народ), пигмееев биака и западноафриканских земледельцев народности мандинка (Сенегал/Гамбия), пришли к выводу, что около 2% генетического материала были получены этими народами от некого ископаемого таксона, обитавшего в Центральной Африке около 35 тыс. лет назад. По их расчётам данный неизвестный вид гоминид отделился от линии, ведущей к современному человеку, примерно 700 тыс. лет назад [18]. Можно отметить, что у представителей вышеупомянутого народа мандинка из Гамбии (индивиды HG02645 [29] и HG02666 [30]) обнаружены дочерняя для А0-Т гаплогруппа A1-V168 (сформировалась 161300 (169900—152900) лет назад, TMRCA 133400 (139000—127800) лет назад [41]) и дочерняя для А1 гаплогруппа А1а M31 (id: HG02666) [17, Table S1]. Установлено, что у мандинка гаплогруппа А1а-М31 (сформировалась 133400 (139000—127800) лет назад, TMRCA 9900 (11500—8500) лет назад) [42]) встречается с частотой 5% [11].
Durvasula, Sankararaman (2020), изучив данные по полному секвенированию геномов 405 человек из популяций Западной Африки (йоруба (Yoruba) из Нигерии, эсан (Esan) из Южной Нигерии, менде (Mende) из Съерра-Леоне, гамбийцы из западных округов Гамбии), пришли к выводу, что их предки получили от 2 до 19% ДНК от гипотетической «призрачной» (ghost) древней популяции, отделившейся от предков неандертальцев, денисовцев и современных людей до их разделения («Our analyses document introgression in four present-day West African populations from an archaic population that likely diverged before the split of modern humans and the ancestors of Neanderthals and Denisovans»). Примерно 6,6% и 7,0% геномов йоруба и менде несли метки древнего происхождения. По расчётам генетиков, эта архаическая популяция отделилась от предка неандертальцев и современных людей от 360 тыс. лет назад до 1,02 млн. лет назад и впоследствии интрогрессировала в предков современных африканцев от 0 до 124 тыс. лет назад. Привлечение геномов неафриканских популяций (китайцы Пекина и жители штата Юта, имеющие северо- и западноевропейское происхождение) показало, что вклад ghost есть и у них. Это позволяет предположить, что скрещивание современного человека с ghost произошло до разделения африканской и неафриканской популяций [15].
В связи с приведёнными выше датами от 360 тыс. лет назад до 1,02 млн. лет назад, отметим, что расчёты с помощью программного обеспечения BEAST2 показали, что время жизни последнего общего предка современных людей и неандертальцев (TMRCA) ~408 тыс. лет назад [16]. Соответственно, «призрачная» (ghost) древняя популяция должна была отделиться от общих предков современного человека и неандертальцев ранее 408 тыс. лет назад.
Если исходить из возраста Y-хромосомных гаплогрупп, то на роль популяции, которая скрещивалась с «призрачной» (ghost) древней популяцией, подходят носители гаплогруппы A1b (сформировалась 133400 (139000—127800) лет назад, TMRCA 130700 (136400—125100) лет назад), материнской для A1b1, распространённой, в основном в Африке и ВТ, по большей части, внеафриканской. В меньшей степени на эту роль подходит популяция носителей материнской для A1b клады A1 (сформировалась 161300 (169900—152900) лет назад, TMRCA 133400 (139000—127800) лет назад) [41].
Xu et al. (2017) изучая белок муцин-7, исследовали 251 геном случайно выбранные из представителей современных европейских, африканских (эсан из Нигерии, жители Западной Гамбии, лухья (Luhya) из Кении, менде из Сьерра-Леоне, йоруба из Нигерии)) и азиатских популяций и установили, что у африканцев, живущих южнее Сахары, ген MUC7, отвечающий за синтез белка муцин-7, резко отличается от обычного для других людей. Более того, MUC7 африканцев отстоит от MUC7 остальных людей намного дальше, чем MUC7 современного человечества в целом отстоит от такого же гена неандертальцев и денисовцев. Данный факт можно объяснить только заимствованием этого гена африканцами у другого вида древних людей. По расчётам Xu et al. (2017) африканский MUC7 был получен людьми, жившими к югу от Сахары, 150 тыс. лет назад, т.е. в это время произошла гибридизация предков современных африканцев с архаичным видом человека, отделившимся от линии, давшей Homo sapiens sapiens 1,5–2 млн лет назад [40].
Дата, полученная Xu et al. (2017), 150 тыс. лет назад, превышает возраст гаплогруппы A1b (133400 (139000—127800) лет назад) и её дочерних клад A1b1 и ВТ (130700 (136400—125100) лет назад). С учётом того, что ВТ распространена, в основном, вне Африки, а у внеафриканского человечества ген MUC7 отличается от африканского, можно полагать, что с древним видом человека скрещивались носители гаплогруппы A1b1.
Исходя из вышеприведённой информации можно сделать вывод, что человек современного анатомического типа скрещивался в Африке как минимум с двумя видами архаичных людей (если верны датировки Hammer et al. (2011) [18], то с тремя видами): ~130 тыс. лет назад предки африканцев и внеафриканских популяций, возможно, носители гаплогрупп A1 и A1b, скрещивались с какой-то «призрачной» (ghost) древней популяцией и ~130 тыс. лет назад предки современных африканцев, возможно, носители гаплогруппы A1b1 (сформировалась 130700 (136400—125100) лет назад [41]) скрещивались с каким-то архаичным видом человека, возможно, с той «призрачной» (ghost) древней популяцией.
Можно также предположить, что в Африке актов гибридизации отдельных популяций человека современного анатомического типа с древними видами людей было больше.
3.2. Археологические культуры/индустрии, создатели которых могли скрещиваться с человеком современного анатомического типа
3.2.1. Ашело-ябрудийский культурный комплекс Леванта (АЯКК)
Ашело-ябрудийский культурный комплекс (АЯКК) включает три основные индустрии: ашело-ябрудьен, ябрудьен и амудьен, развитие которых укладывается в интервал 420–200 тыс. лет назад [6, с. 44].
Ашело-ябрудийские комплексы известны в скальных убежищах Ябруд I, Табун Е, Мислия, Кесем, Зуттие, Хайоним, Абри Зумоффен, Маслоух, Джефр Аджла, Дедерия и др., а также на стоянках под открытым небом, в основном, в центральной и южной частях Леванта [6, с. 46]. В пещере Табун ашело-ябрудийская индустрия имеет следующие даты: слои Ed–Ea — 385–240 тыс. лет назад, нижний слой Ed — 387±49–36 тыс. лет назад (метод сопряжённого электронного спинового резонанса и датирования по урановому ряду (ESR/U-series), слой Е — 260,0±60, 270±60, 340±80, 410±110 и 480±120 тыс. лет назад [2, с. 20].
Пещера Мугарет-эль-Зуттие
Пещера Мугарет-эль-Зуттие находится в Израиле, в Восточной Галилее, на северо-западном берегу Тивериадского озера (=Галилейское море =Генисаретское озеро), около Табги, на левом берегу реки Вади-Эль-Амуд (Wadi Amud). В пещере вместе с артефактами ашело-ябрудийского комплекса (350–300 тыс. лет назад) найдены останки (лобная, правая скуловая и частично сохранившаяся клиновидная кости) гоминина, названного Зуттиех или Галилейский человек. Ашело-ябрудийские артефакты позволили отнести его к архаичной популяции, населявшей Африку, т.е. к таксону, которому принадлежат находки Бодо, Эландсфонтейн, Броукен-Хилл, Эяси, Ндуту. Зуттиех ассоциировали также с ранней архаичной группой Homo sapiens, из-за особенностей лобной кости сближали с ранними неандертальцами и считали прямыми предками людей из Схул и Кафзех [3, с23].
В БРЭ отмечается, что обнаруженные «фрагменты черепа с наклонным лбом, массивным надглазничным валиком, толстой костью свода — древнейшая находка человека в Западной Азии (т. н. Галилейский человек). Первоначально был отнесён к неандертальцам. Позднее было замечено сочетание эректоидных (сходство с пекинским синантропом) и сапиентных признаков (сходство с черепом Кафзех 6), дающее основания причислить его к гейдельбергскому человеку» [8]. Исследователи также отмечают морфологическое сходство Зуттиех и загросских неандертальцев (Шанидар V), среднеплейстоценовых гомининов (Араго XXI) и ближневосточных ранних людей современного типа (Схул V) и приходят к выводу, что «морфология Зуттиех типична для популяции, являвшейся предковой для неандертальцев и людей современного типа или же популяции, существовавшей сразу после расхождения этих двух видов» [3, с. 23].
Кесем (Qesem)
Пещера Кесем расположена в Израиле, в 12 км к востоку от г. Тель-Авива на западном подножии холмов Самарин. Здесь обнаружен археологический материал, характеризующий финальный этап в развитии амудийской индустриальной традиции [2, с. 16].
В пещере Кесем в хронологическом интервале 420–200 тыс. лет назад доминирует амудийская традиция с хорошо выраженными пластинчатыми технологиями. Для ябрудийской индустрии характерны скрёбла кина и полукина и относительно небольшая доля пластин. В обоих комплексах представлены редкие рубила. «В целом амудийская и ябрудийская индустрии из этой пещеры являются частью одной и той же технологической системы, демонстрируя различия главным образом в количественном соотношении категорий инвентаря (например, пластин, орудий на пластинах, скрёбел), т. е. обе индустрии включают скрёбла кина и полукина, а также пластинчатое производство, но в разных пропорциях. Такая вариабельность обусловлена не сменой носителей культурных традиций, а изменениями в хозяйственном укладе и адаптационных стратегиях обитателей пещеры, пространственной и временной дифференциацией деятельности человека на разных участках стоянки. Данные геохронологии свидетельствуют о сосуществовании в пещере Кесем амудийской и ябрудийской индустрий» [6, с. 45].
«Трасологический анализ представительной выборки скрёбел из пещеры Кесем показал, что изделия кина применялись в основном для обработки шкур скоблением, а также твёрдых и среднетвёрдых материалов, в то время как скрёбла полукина были задействованы для резки сырья растительного происхождения и мягких тканей животных [6, с. 48].
Деревянко (2016) отмечает, что в орудийном наборе пещеры Кесем наиболее выразительными являются пластинчатые изделия. При этом, доля пластин в культуросодержащих горизонтах пещеры возрастала снизу вверх, а количество отщепов, используемых в качестве заготовок, снижалось [2, с. 16, 21].
В целом, пластины характерны для амудийского индустриального комплекса, играя в нём важную роль. «Из пластин изготавливали скребки, скрёбла, резцы, зубчато-выемчатые изделия и другие инструменты» [2, с. 17]. При этом, пластинчатые сколы, называемые ножами с естественным обушком, редко подвергались ретушированию [2, с. 17].
Пластинчатая технология впервые появилась в Восточной Африке, в формации Каптурин (Кения), где зафиксирована в культуросодержащих слоях, датируемых 545±3 и 509±9 тыс. лет назад вместе с останками Homo rhodesiensis [2, с. 16; 3, с. 20]. В формации Каптурин пластинчатая индустрия известна также в позднеашельский период, около 280 тыс. лет назад, но она отличается от залегавшей в более древних отложениях и не имеет никаких аналогий с ашело-ябрудийской индустрией Леванта [3, с. 20].
В пещере Кесем обнаружены свидетельства использования костяных фрагментов для вторичной обработки каменных заготовок. Костяной ретушер из израильской пещеры является древнейшим таким орудием в Средиземноморье» [6, с. 50]. Здесь также «помимо обожжённых костей со следами порезов и раскалывания зафиксированы долговременные очаги. Способы разделки туш животных обитателями пещеры Кесем были довольно специфическими и отличались от стратегий мясозаготовки у ашельских, средне- и верхнепалеолитических охотников-собирателей этого региона» [6, с. 50].
В пещере Кесем вместе с каменными артефактами, относящимися к амудийской индустрии, найдены человеческие останки, представленные зубами. Первоначально, Hershkovitz et al. (2011), изучив морфологию зубов из Кесем, выдвинули гипотезу, что обитатели пещеры относятся к местной архаичной популяции Homo, жившей в Юго-Западной Азии 400–200 тыс. лет назад, в то же время, зубы людей из пещеры Кесем, несмотря на некоторую плезиоморфность, указывают на бóльшую степень их родства с популяциями Схула и Кафзеха, нежели с неандертальцами. В пользу гипотезы об их местном происхождении, по мнению исследователей, свидетельствуют археологические материалы: леваллуазские комплексы с большим количеством заготовок в виде пластин и орудий, оформленных на пластинах, подтверждающие местные истоки амудийской индустрии [21]. Согласно недавнему исследованию Hershkovitz et al. (2021), обитатели пещеры Кесем, судя по зубам, имеют определённое сходство с людьми из Нешер Рамла, с фрагментами женского скелета из пещеры Табун (фация С), и фрагментами черепа из местонахождения Мугарет-эль-Зуттие (Zuttiyeh) в Израиле, что позволяет считать этих древних людей представителями отдельной ближневосточной линии людей, отделившейся от неандертальско-денисовского ствола не менее 400 тыс. лет назад [20].
Новосибирские археологи полагают, что часть поздних Homo heidelbergensis, создателей ашело-ябрудийского культурного комплекса Леванта, около 450–350 тыс. лет назад мигрировала с территории Ближнего Востока «в восточном направлении и стала основой для формирования денисовцев, которые достигли юга Сибири и заселили Денисову пещеру на Алтае не позднее морской изотопной стадии 9» [6, с. 43].
Ниже будет показано, что эти древние люди также мигрировали в Африку, где вполне возможно могли скрещиваться с современным человеком.
3.2.2. Сангоан/Лупембан
Более всего на роль архаичного вида людей, скрещивавшегося в Африке с человеком современного анатомического типа, подходят создатели индустрии Сангоан/Лупембан.
Сангоан
Сангоан — археологическая культура эпохи палеолита в Экваториальной и Восточной Африке, на территории Танзании, Кении, Уганды, Замбии и др. [9]. Индустрия сангоан наиболее распространена в лесном поясе Центральной Африки и на территориях вокруг него. Обнаружена она также в Северной, Западной, Южной и Восточной Африке (Симби (Simbi) в Кении) [35].
Названа по стоянкам на холмах Санго в Южной Уганде [9] или в честь залива Санго (Sango Bay) на западном берегу озера Виктория в Уганде. [35].
Несколько датированных стоянок с индустрией сангоана относятся к периоду 250—300 тыс. лет назад [35].
Главный памятник — стоянка Каламбо-Фоллс. Каменные орудия — ручные рубила, колуны, чопперы, свойственные ашельской культуре, многочисленные скрёбла, скребки и зубчатые орудия из отщепов, а также характерные долотовидные рубящие орудия. Сменилась культурой лупембе [9].
В последние годы Taylor (2022), исследовав сангоанскую стоянку Каламбо-Фоллс (Kalambo Falls) в Замбии, пришёл к выводу, что индустрию сангоана сложно отнести как к ESA, так и к MSA [35].
Сангоан сменил ашельскую культуру [9]. Сангоан перекрывает ашель на таких местонахождениях, как Нсонгези (Nsongezi) в Уганде и Каламбо Фоллс (Kalambo Falls) в Замбии [35].
Согласно Taylor (2024), на большей части территории Африки появление сангоана связано с самым ранним технологическим переходом от ашельских рубил (handaxes) к наконечникам (points), пластинам (blades) и более мелким бифасам (smaller bifaces), характерным для MSA [35]. Kuman et al. (2020) утверждают, что переход от ашеля к MSA в Каламбо-Фоллс произошёл примерно 500—300 тыс. лет назад (индустрия санго (Sangoan)). В то время как в ашеле региона подготовленные (prepared) нуклеусы отсутствуют, в сангоане они изредка встречаются в сопровождении редких рубил и кливеров, нуклеусов-рубил (core-axes), нуклеусов-скрёбел (core-scrapers) и редких нуклеусов-пластин (blade cores) [24]. В Каламбо-Фоллс, в слоях санго появляются сколы, характерные для Леваллуа, много крупных скрёбел, мало рубил и кливеров [5].
Clark (1986) датировал сангоан в Каламбо-Фоллс 46—14 тыс. лет до н.э. [14, p.287].
На стратифицированном местонахождении 8-В-11 на острове Сай (Северный Судан) в период перехода от раннего каменного века к среднему (около 200 тыс. лет назад) берега небольшого оврага неоднократно были заселены группами людей, оставившими ашельскую и сангоанскую материальные культуры.
Самый нижний уровень содержит позднеашельскую индустрию с крупными ланцетовидными ручными рубилами. OSL-датирование вышележащих эоловых песков (ES) дало максимальный возраст в 223±19 тыс. лет. Выше этих песков обнаружены три археологических горизонта с каменными орудиями, которые можно квалифицировать как сангоанские. По хронологическому положению они названы Нижний, Средний и Верхний Сангоан. Наиболее характерной типологической особенностью индустрий является наличие тонких бифациальных острий (bifacial foliates), полученных в результате леваллуазской (Levallois), нубийской (Nubian) и дисковидной (discoidal) стратегий расщепления. С технологической точки зрения они относятся к раннему верхнеплейстоценовому нубийскому комплексу, имеющему сходство с лупембанским, расположенным к югу от Сахары [38].
В отличие от ашеля, в ранних уровнях сангоа MSA зафиксировано сложное поведение, требующее значительных технологических и символических знаний. В частности, известны комплексная переработка растительных продуктов (кварцитовые гальки использовались при измельчении растительного сырья) и использование пигментов, свидетельствующих о символической деятельности (жёлтую и красную охру добывали и измельчали до пигментов). В Восточной Африке переход ESA/MSA произошёл более 285 тыс. лет назад. Это может быть свидетельством того, что появление среднего каменного века в Северной Африке является следствием замещения населения [38; 39, р. 9].
Таксономический статус создателей индустрии Сангоан
На озере Эяси (Северная Танзания), вместе с каменными орудиями, относящимися к сангоану и датированными> 130 тыс. лет назад, найдены человеческие окаменелости [26].
Bosman et al. (2020) исследовали череп Eyasi I (озеро Эяси, Танзания) (375–88 тыс. лет назад), у которого в более ранних исследованиях была обнаружена надбровная ямка (suprainiac fossa), углубление на затылочной кости над затылочным бугром, которая считается аутапоморфией (признаком) неандертальцев. Они установили, что ни внешняя, ни внутренняя морфология этих углублений не имеют никакого сходства с неандертальской. Анализ главных компонентов, основанный на многократных измерениях толщины затылочной кости, показывает, что относительные значения толщины внутренних структур у Eyasi I находятся в пределах диапазона Homo sapiens [12].
Череп Homo helmei Синги из Абухагара (Abu Hagar; Восточный Судан, западный берег Голубого Нила, 320 км юго-восточнее Хартума) найден с каменными орудиями, и, вероятно, в той же стратиграфической позиции, что и индустрия сангоан на острове Сай (Sai Sangoan) [38]. На сайте Смитсоновского Национального музея естественной истории (Вашингтон, США) Синга 1, возрастом между 150 и 120 тыс. лет назад, опредедяется как Homo sapiens [31].
Люпембан
Лупембан — индустрия среднего каменного века (MSA) в Центральной и Западной Африке (на территории Анголы, Конго, Заира), получившая своё название от местонаждения Лупемба (Lupemba) в Конго (в прошлом — Бельгийское Конго). Большая часть материалов — сборы с поверхности стоянок. Распространены как архаичные орудия из камня (рубила, чопперы, колуны), так и орудия с развитыми формами (крупные листовидные острия, долотовидные орудия). Много пестов-тёрочников и плит для растирания продуктов собирательства [5; 7].
Существуют разные точки зрения на соотношение сангоана и лупембана, от люпембан — культура, родственная санго [5], до лупембан сменил сангоан [9].
На основе культуры лупембан развивается читольская культура Центральной Африки [7], известная, в первую очередь, в Заире, а также в регионе Лунда (Lunda region) на северо-востоке Анголы. Она распространена в нижнем течении реки Конго и в бассейне её южных притоков, таких как Касаи и Кванго, от среднего до верхнего течения. Также встречается за водоразделом реки Конго на плато, особенно к югу и востоку от её бассейна [27].
Taylor (2022) утверждает, что большие двусторонние ланцетовидные наконечники (large bifacial lanceolate points), характерные для местонахождений люпембана Каламбо Фоллс, Твин Риверс, Мугурук, отсутствуют в сангоане. Вместе с тем, большие нуклеусы-рубила (heavy-duty core-axes) и кирки (picks) люпембана предполагают его родственные связи с сангоаном и поздним ашелем, в которых они также присутствуют [34]. По его мнению, в широком смысле, люпембан, по-видимому, является свидетельством первоначального и широкого распространения в Центральной Африке технологии подготовленного нуклеуса (prepared core technology (PCT)), пластин (blades), а на некоторых участках — пластин с геометрической спинкой (geometric backed blades). В Твин-Риверс также наблюдается большое количество антропогенно накопленных пигментов [34].
Вместе с тем, можно предположить разное происхождение сангоана и лупембана. Так, например, Van Peer et al. (2004) утверждают, что в Саи 8-В-11 на острове Сай (Северный Судан) два самых нижних слоя можно отнести к сангоану из-за наличия нуклеусов-рубил (core-axes) и стратегий расщепления отщепов (flake reduction strategies). С учётом наличия систематического производства пластин (blade) и мелких рубил ланцетовидной формы верхний комплекс этой последовательности квалифицируется как лупембан. Нуклеусы-рубила Саи 8-B-11 так называемого типа Хор-Абу-Анга (Khor-Abu-Anga) сопоставимы с лупембанские нуклеусами-рубилами Каламбо Фоллс [39, р.9].
По мнению Van Peer et al. (2004) к лупембану нужно относить ассамбляжи MSA, где представлены стратегии расщепления пластин (blade), а к сангоану индустрии, в которых применяются стратегии отщепов (flake) и представлены листовидные орудия (foliates) в примерно одинаковых пропорциях с нуклеусами-рубилами (core-axes) [39, р.12].
Согласно результатам исследований, территория распространения люпембана совпадает с современным поясом тропических лесов Конго. На окраинах ареала распространения люпембана находятся местонахождения Каламбо Фоллс (Kalambo Falls), Твин Риверс (Twin Rivers), Мугурук (Muguruk).
На большей части географического ареала индустрии люпембана датированы плохо. На местонахождении в Твин-Риверс в Замбии получена серия урановых дат ~270–170 тыс. лет назад [34].
Taylor (2022) предполагает, исходя из дат Твин-Риверс, что люпембан возник на окраинах Центральной Африки примерно 270 тыс. лет назад и продолжал существовать до 170 тыс. лет назад. Набор инструментов лупембана, состоящий из удлинённых ланцетовидных бифасов, нуклеусов-топоров (core-axes), острий (points), лезвий (blades) и других инструментов, характерен для пояса экваториальных лесов и по мнению исследователей, появился в MIS 7 в результате адаптации к добыче пищи в тропическом лесу. После того, как в MIS 6 тропические леса начали сокращаться, создатели инструментов Лупембана, возможно, расселились по равнинной части Центральной Африки в течение сотен или даже тысяч человеческих поколений [34].
Существует предположение, что орудия люпембана, распространённые в современном лесном поясе Конго, могли быть приспособлены для обработки дерева. Обоюдоострые ланцетовидные наконечники интерпретируются как сохранившиеся элементы составных копий [13]. Короткие копья пигмеев мбути в лесу Итури имели подобные по форме железные лезвия (около 30×20 см), укреплённые на короткой (около 75 см) и толстой рукоятке из твёрдой древесины. С такими копьями они охотились на слонов (пробирались под стоящего слона и молниеносным движением вонзали копьё в его мягкое брюхо) и кабанов [34].
Miller (2001) отмечает сходство некоторых элементов культуры (плетение корзин) читолиана (Tshitolian), археологической культуры, считающейся наследницей люпембана и баяка (Bayaka) в провинции Кванго в Заире [27, p. 282], пигмеев, известных также как ака (Aka), биака (Biaka), бабензиле (Babenzele). Охотники-собиратели Южной, Центральной и Восточной Африки (сан (San), кой-коин (Khoikhoi; khaa, khaasen) [готтентоты], пигмеи (Pygmäen) и др.) до недавнего времени использовали орудия лупембан-читольского (Lupembo-Tshitolian) типа [22].
Здесь нужно вспомнить, что Hammer et al. (2011) нашли у пигмеев биака около 2% генетического материала, полученного ими от некого ископаемого таксона, обитавшего в Центральной Африке около 35 тыс. лет назад [18].
Таксономический статус создателей индустрии лупембан
Можно было бы предположить, что создатели индустрии лупембан относятся к виду Homo heidelbergensis, поскольку гоминид из пещеры Мумбва (Mumbwa), расположенной неподалёку от местонахождения Твин-Риверс, индустрия люпембан которого датируется 270—170 тыс. лет назад [34], был классифицирован как Homo heidelbergensis [1, с.84]. В частности, согласно Pearson et al. (2000), зубы, происходящие из уровней MSA (Mumbwa E10–1–0) в пещере Мумбва, за небольшими исключениями, показывают сходство с зубами современных жителей Африки к югу от Сахары. В то же время, посткраниальные останки (бедренная кость) из Мумбвы, приписываемые MSA (Mumbwa E10–1–0) показывают сходство с таковыми среднеплейстоценовых Кабве [Брокен-Хилл, Кабве / Broken Hill, Kabwe; Родезийский человек / Rhodesian Man; Homo heidelbergensis] и KNM-ER 999, но принадлежат особи гораздо меньшего размера, чем экземпляры KNM-ER 999 и Kabwe. Посткраниальные останки E9–3–24 MSA из Мумбвы по такому признаку как радиальная бугристость проксимального фрагмента, близки к таковой для большинства современных людей, а также Homo antecessor, Homo heidelbergensis и раннего Homo neanderthalensis из Атапуэрки и отличаются от неандертальцев [28].
Результаты исследования Steyn et al. (2023) показали, что найденные в пещере Мумбва плохо сохранившиеся останки (в основном, череп и зубы) принадлежат, как минимум 16 человекам, похороненным здесь где-то между концом X и началом XX века нашей эры, причём большинство дат приходится на начало XVI и конец XIX веков [33]. Поэтому таксономический статус создателей индустрии лупембан пока не ясен.
Поскольку лупембан датируется ~270—170 тыс. лет назад и предположительно возник на окраинах Центральной Африки в тропических лесах [34], то вряд ли создатели этой индустрии были людьми современного анатомического типа. Жизнь в тропическом лесу предполагает специализацию, ведущую к образованию отдельного вида. Современный человек мог сформироваться лишь в зоне с разнообразными природно-климатическими условиями, требующими постоянной адаптации к изменяющимся условиям окружающей среды и соответственно, приводящих к развитию умственных способностей. Такие разнообразные условия имелись в Восточно-Африканской рифтовой долине, где и происходила эволюция современного человека.
Поскольку есть гипотезы (или теории) о том, что современный человек скрещивался с какой-то «призрачной» (ghost) древней популяцией до разделения африканской и неафриканской линий и которая интрогрессировала в предков современных африканцев до 124 тыс. лет назад [15], а также, что предки современных африканцев скрещивались с каким-то архаичным видом человека 150 тыс. лет назад [40], можно предположить, что создатели индустрии лупембан — это и есть «призрачная» (ghost) древняя популяция.
Каламбо-Фоллс
По мнению Taylor (2022) Каламбо Фоллс — единственный лупембанский комплекс в изолированном стратиграфическом контексте и, таким образом, представляющий собой единственный однозначно лупембанский комплекс, известный в настоящее время [34].
В Каламбо-Фоллс выделены три хронологические стадии в развитии индустрии люпембан: ранняя, накисаса; средняя, сисция; поздняя, полунг [4, с. 88].
В индустрии раннего и среднего люпембана на этой стоянке присутствуют двусторонние копьевидные острия, орудия типа бифасов, нуклевидных топориков, а также более мелкие изделия: скрёбла, скребки, проколки и др. Отсутствуют обушковые орудия геометрической формы [4, с. 89]. Деревянко (2020) предполагает, что их отсутствие, возможно, объясняется действием водных потоков, которые «вымыли» небольшие по размерам каменные изделия. В первичном расщеплении выделяются леваллуазские и пластинчатые нуклеусы [4, с. 89].
В среднем люпембане Каламбо-Фолс исследователи отмечают присутствие усечённых отщепов и пластин, обушковых изделий геометрической формы. В культуросодержащих горизонтах среднего люпембана с индустрией сисция, как и в Твин Риверс, зафиксировано систематическое использование гомининами пигментов [4, с. 90].
Мугурук
Стоянка Мугурук расположена на восточном берегу реки Мугурук, в 3 км к северу от берега залива Винам (Winam Gulf) озера Виктория, на западе Кении, на окраине ареала распространения индустрии люпембан [34].
В Мугуруке обнаружены комплексы, относящиеся к переходному периоду Сангоан-Лупембан/MSA. Ассамбляжи Мугурука относятся к более ранней индустрии Оджолла (Ojolla) и более поздней индустрии Пундо Маквар (Pundo Makwar). McBrearty (1988) артефакты индустрии Оджолла (Ojolla) относит к Сангоан-Лупембану (Sangoan‐Lupemban), а индустрию Пундо Маквар (Pundo Makwar) к среднему каменному веку (MSA) [25].
Taylor (2022) полагает, что в Мугуруке смешанная сангоан-лупембанская (Оджолла) индустрия с ланцетовидными инструментами, перекрывается горизонтом недиагностируемого материала MSA [34].
По McBrearty (1988) индустрия Оджолла [т.е. сангоан-лупембан] включает в себя тяжёлые режущие инструменты, тонко обработанные ланцетовидные наконечники и лёгкие компоненты, состоящие из двусторонне ретушированных сколов отщепов (flake tools). Производство отщепов осуществлялось путём радиального расщепления нуклеусов. Индустрия Пундо Маквар состоит в основном из отщепов, образующихся в результате радиального расщепления нуклеусов и односторонних скребков, изготовленных на этих отщепах. По её мнению, археологические материалы Мугурука демонстрируют выраженный сдвиг в методах ретуши инструментов между Сангоан-Лупембаном и средним каменным веком, но реальную преемственность в методе производства отщепов [25].
Согласно Taylor (2022), определяющими для ассамбляжа Мугурука являются обоюдоострые ланцетовидные наконечники [34].
Shea (2008) сравнивает индустрию Мугурука с индустрией Омо Кибиша, где обнаружены останки древнейшего человека современного анатомического типа и констатирует, что индустрия Оджолла [т.е. сангоан-лупембан] существенно отличается от индустрии Кибиша тем, что в ней используются тяжёлые двусторонние режущие инструменты и относительно редки леваллуазские материалы. Индустрия Пундо Маквар [т. е. MSA] имеет многое из ключевых особенностей индустрии Омо Кибиш — выраженный леваллуазский компонент, относительно небольшое количество бифасиальных отщепов, мелкие листовидные бифасы и преимущественно радиальная/центростремительная подготовка нуклеусов [32].
Индустрия стоянки Мугурук не датирована. У Tryon, Tyler Faith (2013) Мугурук с индустрией Пундо Маквар отнесены к LMSA? (поздний африканский средний палеолит). Возраст указан> 10–170 тыс. лет назад. Такой же возраст указан для более древнего Мугурук с индустрией Оджолла — EMSA? (ранний африканский средний палеолит) (Неопределённость, по их мнению, возникает для недатированных участков, которые находятся на границе ~ 126 тыс. лет назад, между EMSA/LMSA, или где биостратиграфические данные отличаются от радиометрических оценок возраста) [37].
IV. Гаплогруппа А00
4.1. Общая характеристика
Гаплогруппа А00 (рис. 14) cформировалась 235900 (243700—228300) лет назад, TMRCA 37600 (42200—33100) лет назад. Базальная линия этой гаплогруппы обнаружена у индивида возрастом 7885 лет назад в скальном убежище Шум Лака (Shum Laka) (рис. 15) на северо-западе Камеруна [15]. Человек с местонахождения Шум Лака: I10871 — Y-DNA — A-L1087 (A00), mtDNA — L0a2a1 [5].
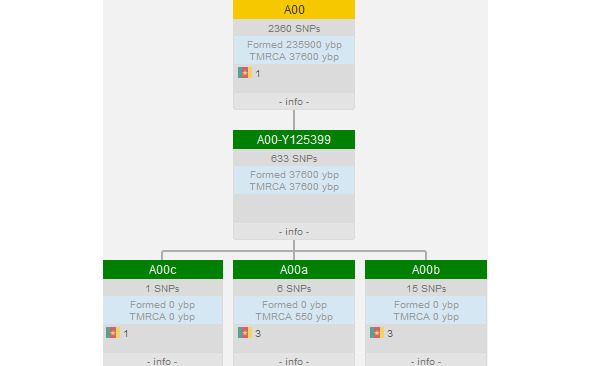
Рис. 14. «Древо» гаплогруппы А00 [31]
В настоящее время гаплогруппа А00 встречается только в Камеруне [15]. Дочерние для А00 клады обнаружены со следующими частотами: A00a (L1149, FGC25576, FGC26292, FGC2693, FGC27741) — у бантуязычных бангва и мбо — 40% и 6,3%, соответственно (Bangwa of Cameroon (40%) & Mbo of Cameroon (6,3%)); A00b (A4987/YP3669, A4981, A4982/YP2683, A4984/YP2995, A4985/YP3292, A4986, A4988/YP3731) — у мбо — 9,3% (Mbo of Cameroon (9,3%)) [4]. Носитель гаплогруппы А00с также обнаружен в Камеруне [31].
В вышеупомянутой Шум Лака наиболее древние следы обитания датируются ~30 тыс. лет назад [15]. Примечательно, что возраст следов обитания в Шум Лака примерно совпадает со времем жизни последнего общего предка современных представителей гаплогруппы А00 (TMRCA 37600 (42200—33100) лет назад [31]). Это дата может быть свидетельством миграции в Камерун носителей гаплогруппы А00 из какого-то другого региона.
Рис. 15. Шум-Лака на карте Камеруна [9]
Поскольку у древнего представителя с Y-ДНК A00 определена мтДНК L0a2a1, можно предположить, что древнейшая популяция носителей Y-ДНК A00 была также носителем мтДНК L0.
4.2. ДНК древних обитателей местнахождения Шум Лака. Гипотеза Lipson et al. (2020)
Lipson et al. (2020) секвенировали полногеномные данные ДНК четырёх человек, похороненных в Шум Лака — мальчиков 2/SE I (I10872) и 2/SE II (I10871) в возрасте 4±1 и 15±3 лет, соответственно, датированных около 8 тыс. лет назад и 8±2-летнего мальчика 4/A (I10873) и 4±1-летней девочки 5/A, датированных 3 тыс. лет назад [15]. У мальчика 2/SE I определили митохондриальную гаплогруппу L0a2a1 и Y-хромосомную гаплогруппу В, характерные для современного населения Африки южнее Сахары. У мальчика 2/SE II определены митохондриальная гаплогруппа L0a2a1 и древнейшая Y-хромосомная гаплогруппа A00, имеющая возраст ~235900 тыс. лет [15, Table 1]. Повторный анализ Y-хромосомы обитателя пещеры Шум Лака, возрастом ~8 тыс. лет назад, показал, что он относится к линии A00b [17].
У детей, датированных 3 тыс. лет назад определена митохондриальная гаплогруппа L1c2a1b, распространённая среди населения Центральной и Западной Африки. Мальчик принадлежит к Y-хромосомной гаплогруппе B2b [15, Table 1]. Повторное переопределение гаплогруппы индивида из Шум Лаки, у которого была первоначально определена B2b-M112, показало, что у него линия B2b1-M192. При этом, он наиболее близок к современному представителю народа лухья (Luhya) из Кении (народ банту и вторая по численности группа в Кении), несущего линию B2b1, а не к представителям народностей биака (ака (Aka), bayaka, babenzele — пигмеи группы западные бамбенга (Bambenga) или мбенга (Mbenga), мбути (восточные пигмеи) и сан (бушмены), носителям гаплогруппы B2-M82 [17].
На сайте компании Y Tree Limited индивид из Шум Лаки I10873 (мальчик 2/SE I) отнесён к сестринской для B-M192 гаплогруппе B-CTS591 (сформировалась 48700 (51800—45800) лет назад, TMRCA 18900 (22400—15800) лет назад и датирован 3086 (3005—3167) лет назад. К этой же гаплогруппе принадлежит индивид NA19384 из Кении [34], представитель народа лухья (Luhya) [22]. Примечательно, что TMRCA гаплогруппы B-CTS591 совпадает с датой разделения пигмеев на западную и восточную ветви — примерно 20 тыс. лет назад (21.9 Kya (95% CI: 14.2–66.3)), согласно Patin et al. (2009) [20].
Использовав метод анализа главных компонент, Lipson et al. (2020) установили, что индивиды из Шум Лаки генетически далеки от народов банту и родственных им западноафриканских популяций (чева (Chewa), мбо (Mbo) и менде (Mende)) и ближе всего к современным охотникам-собирателям/пигмеям (негрилли) из Камеруна (бака (Baka), бакола (Bakola) и бедзан (Bedzan)) и Центральноафриканской республики (ака (Aka) или биака (Biaka)). В то же время современные пиггмеи Камеруна не являются их прямыми потомками [15].
Исходя из вышеизложенных результатов, Lipson et al. (2020) делают далеко идущие выводы. В частности, они выделяют на построенном ими филогенетическом дереве три основных узла ветвления (рис. 16). В первом узле, по их мнению, 300—200 тыс. лет назад, разделяются охотники-собиратели Южной Африки и Центральной Африки. Центральноафриканские охотники-собиратели затем делятся на восточную и западную ветви, от которой впоследствии произошли люди из Шум Лаки (рис 16а).
Рис. 16. Узлы ветвления на филогенетическом дереве [15, Fig. 4].
Во втором узле ветвления (точка 2 на рис. 16b) разделились популяции Западной Африки и Восточной Африки и ветвь, вышедшая из Африки. Исследователи датируют это событие — 80—60 тыс. лет назад. По их мнению, такая датировка согласуется с филогеографией мтДНК — в частности, с диверсификацией гаплогруппы L3, которая, по их мнению, вероятно, возникла в Восточной Африке около70 тыс. лет назад и, возможно, с происхождением клады CT на Y-хромосомном дереве на аналогичной временной глубине.
Третий узел ветвления (точка 3 на рис. 16b) связан с выделением бантуязычных популяций [15, Fig. 4].
У Lipson et al. (2020) есть информация о каменных орудиях, обнаруженных в Шум Лаке — микролитическая кварцевая индустрия (microlithic quartz industry), макролитические отщепы и пластины из базальта (macrolithic flake and blade industry on basalt), бифасы типа топор-мотыга (bifaces of the axe-hoe type) [15]. К сожалению, нет информации о применявшихся системах расщепления камня.
Поскольку Lipson et al. (2020) установили, что древние жители Шум Лаки наиболее близки современным пигмеям и построили гипотезу о выходе предков современного неафриканского населения из Африки 80—60 тыс. лет назад, затронем тему пигмеев.
Patin et al. (2009) установили, что отделение пигмеев от других бантуязычных народов, скорее всего, произошло около 60 тыс. лет назад (56 Kya (95% CI: 25.8–130.5)), а разделение пигмеев на западную и восточную группы произошло примерно 20 тыс. лет назад (21.9 Kya (95% CI: 14.2–66.3)) [20]. Результаты исследования Patin et al. (2009) частично подтверждаются исследованием Verdu et al. (2009), которые проанализировали ДНК 604 человек из 9 пигмейских и 12 непигмейских народов, обитающих на территории Камеруна, Габона и Конго. Согласно этому исследованию пигмеи и жители равнин произошли от одного предка 20—120 тыс. лет назад (наиболее вероятное значение — 90 тыс. лет назад), и затем несколько десятков тысяч лет не имели притока генов извне. В период от 30 до 1 тысячи лет назад с наиболее вероятным значением 2800 лет назад, пигмеи разделились на группы, по-видимому, под влиянием народов, занимавшихся сельским хозяйством, которые могли прийти на запад Экваториальной Африки несколько тысяч лет назад [29]. Можно отметить, что дата разделения прародительской популяции пигмеев и жителей равнин — примерно 90 тыс. лет назад, близка к датировкам Y-хромосомной гаплогруппы B — сформировалась 88000 (92500—83600) лет назад, TMRCA 84800 (90300—79400) лет назад [34], которая у пигмеев биака и мбути достигает частоты 35% [3, с. 64].
Поэтому можно полагать, что на запад Африки гаплогруппа В пришла относительно поздно и она была основной гаплогруппой пигмеев. Если ориентироваться на TMRCA клады B-CTS591 — 18900 (22400—15800) лет назад, примерно 20 тыс. лет назад эта гаплогруппа появилась на западе Африки. Если же ориентироваться на возраст индивида I10873 — 3086 (3005—3167) лет назад и информацию о присутствии пигмеев на юге Египта в III тыс. до н.э., то она появилась в этом регионе всего лишь около 4 тыс. лет назад. В Западной Африке пигмеи смешались с пришедшими до них в этот регион представителями гаплогруппы А00, где-то на пути в Западную Африку они могли встретиться с «призрачной» (ghost) популяцией древних людей или же получить эту компоненту от носителей клады А00. Возможны и два источника архаичного генетического материала, поскольку у пигмеев биака около 2% генетического материала, получено от некоего ископаемого таксона, обитавшего в Центральной Африке около 35 тыс. лет назад [12].
Пигмеи упоминаются в древнеегипетских документах конца III тыс. до н.э. (рис. 17). До 1933 г. пигмеи встречались в болотистых местностях Белого Нила на Бахр-эль-Араб и Бахр-эль-Газал [1].
Исходя из вышеприведённой информации можно полагать, что гипотеза Lipson et al. (2020), по крайней мере, в части пигмеев неверна. Пигмеи имеют общее происхождение с бантуязычными народами и ещё в III тыс. до н.э. часть из них проживала неподалеку от Египта. Малый рост пигмеев может быть результатом их гибридизации с малорослым гейдельбергским человеком (выше показано, что их рост в Италии составлял ~150 см).

Рис. 17. Музыканты и танцоры на ТТ38. (Инвентарный номер 30.4.9, Метрополитен-музей, Фонд Роджерса, 1930 г.) (Musicians and dancers on the TT38. (Accession number 30.4.9, Metropolitan Museum of Art, Rogers Fund, 1930): https://www.metmuseum.org/art/collection/search/557727 (Public Domain Image) [11, Fig. 4]
Гипотезе Lipson et al. (2020) также противоречат результаты исследования Skoglund et al. (2017), которые установили, что предки койсанов в прошлом были широко расселены на территории Африки, составляя примерно две трети охотников-собирателей Малави, живших 8100—2500 лет назад и примерно одну треть охотников-собирателей, живших около 1400 лет назад на территории Танзании. По их мнению, расселение фермеров из Западной Африки привело к полному вытеснению местных охотников-собирателей в ряде регионов. Расселение скотоводов, живших 3100 лет назад в Танзании, привело к передаче ими своего генетического материала населению от Северо-Восточной до Южной Африки, в том числе 1200-летним южноафриканским скотоводам [24].
Относительно упомянутой выше митохондриальной гпалогруппы L3, то согласно Maier et al. (2022) она сформировалась 68420 (80457—56405) лет назад, TMRCA 57900 (71427—44575) лет назад [16, Table S3] и соответственно не является ровесницей Y-хромосомной гаплогруппы СТ, которая по расчётам компании YFull имеет возраст 88000 (92500—83600) лет назад, TMRCA 68500 (71800—65200) лет назад [32]. При этом, возраст и TMRCA Y-ДНК СТ практически совпадают с возрастом и TMRCA, материнской для L3 мтДНК L3'4, которая согласно Maier et al. (2022) сформировалась она 83584 (95097—71777) лет назад, TMRCA 68420 (80457—56405) лет назад [16, Table S3]. По-видимому, данное совпадение является свидетельством того, что в Евразию из Африки мигрировала популяция-носителей Y-хромосомной гаплогруппы СТ и митохондриальной гаплогруппы L3'4 и соответственно мтДНК L4 (L4 сформировалась 68420 (80457—56405) лет назад, TMRCA 62191 (74525—49844) лет назад [16, Table S3]) — результат обратной миграции из Азии в Африку.
4.3. Митохондриальная гаплогруппа L0 — «спутница» Y-хромосомной гаплогруппы А00 и возможная связь с ними койсанских народов
Логично предположить, что люди с Y-хромосомной гаплогруппой А00, отделившиеся от базальной популяции людей, были также носителями митохондриальной гаплогруппы L0. Косвенно на это указывает присутствие у древних обитателей Шум Лаки как YDNA A00, так и субклад mtDNA L0.
L0, по расчётам генетиков, сформировалась 143454 (160228—127140) лет назад, TMRCA 112943 (129536—97014) лет назад [16, Table S3]. У Schlebusch et al. (2020) для L0 приведена цифра 144,931 лет назад. При этом, сестринская для неё L1’2’3’4 имеет возраст 152,809 лет назад [23, Fig. S5.7]. На мой взгляд, логичным считать, что возраст обеих митохондриальных гаплогрупп одинаковый.
Выше уже приводилась информация Ingman et al. (2000), согласно которым TMRCA для мтДНК составляет 171500±50000 лет назад [13, p.712] и отмечалось, что дата 171500 лет назад очень близка к времени жизни последнего общего предка современных представителей Y-хромосомной гаплогруппы A0-T (TMRCA) -161300 (169900—152900) лет назад) согласно YFull [32]. При проведении небольшого расчёта: 171000+50000=221000 лет, то получим цифру близкую к возрасту Y-хромосомной гаплогруппы А00 — 235900 (243700—228300) лет назад [31]. Поэтому логично предположить, что возраст mtDNA L0 примерно равен возрасту YDNA А00.

Рис. 18. Койсаны [21]
Y-хромосомная гаплогруппа А00, как и А0-Т, скорее всего, родом из Восточноафриканской рифтовой долины и связана с древнейшей группой, отколовшейся от предковой популяции людей современного анатомического типа — койсанскими народами (рис. 18). У этих народов широко представлена митохондриальная гаплогруппа L0 (рис. 6), которая, предположительно была «спутницей» Y-ДНК А00. При этом, клады митохондриальной гаплогруппы L достигают своего пикового разнообразия в странах Восточноафриканской рифтовой долины — Эфиопии, Кении и Танзании [16, Fig. 4], а L0 достигает наибольшей частоты у койсанских народов Южной Африки. Так, в смешанной выборке народов! Сюнь/Кхоэ (!Xun/Khwe), проживающих в районе Шмидтсдрифт на северо-западе Капской провинции ЮАР, L0 обнаружена с частотой 83% (L0d — 61%, L0k — 22%), у кунг (!Kung) в Ботстване L0 — 100% (L0d — 96%, L0k — 4%) [25]. При этом, у этих народов в мужском генофонде нет YDNA А00. Генофонд Кхоэ: A3b1-M51 — 12%, E3a-M2 — 54%, E3b-M35 — 31%, E*, D — 4%; Кунг: A3b1-M51 — 28%, A* — 8%, B2b-M112 — 8%, E3a-M2 — 39%, E3b-M35 — 11%, E*,D — 6% [25, Fig. 4].
В Танзании, обитают, по-видимому, потомки древних койсанов, осташихся на родине (остальные под давлением других народов мигрировали в Южную Африку). У койсанских народов в Танзании хадза (hadza) L0 — 5% (L0a5%), сандаве (sandawe) L0 — 26% (L0d 5%, L0f — 4%, L0a — 17%). Эти народы проживают на расстоянии 150 км друг от друга. Хадза живут неподалеку от озера Эяси в районе Аруша на северо-востоке Танзании (Lake Eyasi in the Arusha district of north-central Tanzania). Сандаве, в основном, в районе Кондоа к юго-востоку от Аруши (Kondoa district southeast of Arusha) [25].
МтДНК-гаплогруппа L0 есть и других народов Танзании: у проживающего неподалёку от сандаве нигер-кордофанского народа туру (turu) L0 — 24% (L0f — 14%, L0a — 10%), у нило-сахарского народа датога L0 — 5% (L0a 5%), у афроазиатскогго народа бурунге L0 — 58% (L0d — 3%, L0f — 30%, L0a — 25 [25, Fig. 3]. Есть L0 и в соседних с Танзанией странах: у населения Мозамбика L0 — 30% (L0d — 5%, L0a — 25%), у населения Эфиопии L0 — <9% (L0d — <1%, L0f — <1%, L0a — 7%), в смешанной выборке Кения/Судан L0 — 12% (L0d — 1%, L0a -11%) [25, Fig. 3.].
В мужском генофонде ни хадза, ни сандаве не обнаружена гаплогруппа А00 (YDNA: хадза B2b-M112 — 52%, E3a-M2 — 30%, E3b-M35 — 13%, C,F-R — 4%; сандаве A3b2-M13 — 4%, B2a-M150 — 1%, B2b-M112 — 13%, E3a-M2 — 43%, E3b-M35 — 34%, E*,D — 5% [25, Fig. 4]).
Учёные предполагают, что охотники-собиратели, предки хадза и сандаве проживали в регионе Танзании, задолго до прибытия сюда популяций, с производящим хозяйством. Скотоводы из Эфиопии, говорящие на кушитских (афро-азиатских) языках впервые достигли Северной Танзании около 4 тыс. лет назад. Вслед за ними из Южного Судана пришли носители нилотских (нило-сахарских) языков. Около 2,5 тыс. лет назад из Западной Африки в Северо-Западную Танзанию пришли народы банту (нигер-кордофанцы) [25]. Поэтому вполне могло произойти замещение А00 в генофонде хадза и сандаве на другие гаплогруппы (В, Е и др.) с приходом в Танзанию нового населения.
Можно проверить правы ли исследователи, утверждающие вышенаписанное. Для этого можно сравнить возраст Y-хромосомных гаплогрупп A3b2-M13, B2b-M112 и Е хадза и сандаве с вышеприведёнными датами миграций различных групп населения в Танзанию.
Гаплогруппа A-M13 сформировалась 44100 (47500—40800) лет назад, TMRCA 10900 (12300—95000) лет назад. Носитель базальной линии одной из её дочерних клад A-YP4751 (сформировалась 10900 (12300—95000) лет назад, TMRCA 10900 (12300—95000) лет назад) обнаружен в Кении (Рифтовая долина). Представители нижележащих линий этой гаплогруппы живут на Аравийском полуострове, в Судане, Чаде, Уганде, Кении, Эфиопии [33]. Интересно, что с датой прихода в Танзанию около 4 тыс. лет назад скотоводов из Эфиопии, разговаривающих на кушитских (афро-азиатских) языках, совпадает возраст гаплогруппы A-BY16295 (сформировалась 3500 (4400—2700) лет назад, TMRCA 3500 (4400—2700) лет назад), представители которой сейчас обнаружены в Судане (A-FGC71697 (сформировалась 3500 (4400—2700) лет назад, TMRCA 50 (200—50) лет назад) и Сомали (A-Y142085 ((4400—2700) лет назад, TMRCA 3400 (4800—2200) лет назад) (носители базальной линии этой гаплогруппы живут в Сомали и Палестине)). Представители вышележащих для A-BY16295 клад живут, в основном, на Аравийском полуострове, но часть из них в Эфиопии (A-Y92096 (сформировалась 4900 (5800—4000) лет назад, TMRCA 4300 (5200—3400) лет назад); A-Y161004 (сформировалась 5100 (6200—4000) лет назад, TMRCA 2400 (3200—1650) лет назад). Родительская для этой группы Y-хромосомных гаплогрупп клада A-Y23865 сформировалась 9800 (11000—8700) лет назад, TMRCA 8300 (9400—7200) лет назад. Она «внучка» гаплогруппы A-M13 [33].
Таким образом, можно констатировать — генетические данные подтверждают мнение исследователей о приходе носителей гаплогруппы A-M13 в Танзанию из Эфиопии около 4 тыс. лет назад.
Гаплогруппа B-M112 сформировалась 58400 (61500—55300) лет назад, TMRCA 51900 (55000—48900) лет назад. В Камеруне живут представители базальной линии дочерней для неё гаплогруппы B-V3220 (сформировалась 51900 (55000—48900) лет назад, TMRCA 6500 (7900—5200) лет назад), а носители дочерней для B-V3220 гаплогруппы B-SK522 живут в Центральноафриканской республике. Также в Кении и Камеруне обнаружены представители второй дочерней для B-M112 гаплогруппы B-Y18023 (сформировалась 51900 лет назад, TMRCA 48700 лет назад), конкретнее дочерней для B-Y18023 гаплогруппы B-CTS591 (сформировалась 48700 (51800—45800) лет назад, TMRCA 19000 (22500—15900) лет назад [34]). Представитель этой гаплогруппы в Камеруне — древний обитатель Шум Лаки I10873 (мальчик 2/SE I), возрастом 3086 (3005—3167) лет назад (у него митохондриальная гаплогруппа L0a2a1), а индивид из Кении [34], представитель народа лухья (Luhya) [22]. При этом, у индивида (2/SE II), похороненного в Шум Лаке 8 тыс. лет назад, определена Y-хромосомная гаплогруппа А00 [15, Table 1], A00b [17]. У него также, как и у жившего на 5 тыс. лет позже вышеупомянутого носителя Y-хромосомной гаплогруппы B-CTS591 определена митохондриальная гаплогруппа L0a2a1 [15, Table 1].
Данный факт может быть свидетельством замещения в популяции, обитавшей в районе Шум Лаки, мужского генофонда в результате прихода из Восточной Африки нового населения.
Представители второй дочерней клады B-Y18023 гаплогруппы B-M192 (сформировалась 48700 (51800—45800) лет назад, TMRCA 44900 (47800—42000) лет назад), живут на Аравийском полуострове, в Конго и Намибии [34].
Миграцию около 2,5 тыс. лет назад из Западной Африки в Северо-Западную Танзанию народов банту (нигер-кордофанцы) можно связать с носителями гаплогруппы B-SK522, представители которой обнаружены в ЦАР, т.е. к западу от Танзании (гаплогруппа сформировалась 6500 (7900—5200) лет назад, TMRCA 1000 (1650—550) лет назад)) [34].
Гаплогруппа B-M150 сестринская гаплогруппа для B-M112. Гаплогруппа B-M150 сформировалась 58400 (61500—55300) лет назад, TMRCA 37500 (40200—34900) лет назад. Представители одной из её дочерних линий B-M6057 (сформировалась 37500 (40200—34900) лет назад, TMRCA 26300 (29000—23700) лет назад) живут в Конго и на Аравийском полуострове. Представители другой её дочерней клады B-V75 (сформировалась 37500 (40200—34900) лет назад, TMRCA 28300 (30900—25900) лет назад) живут на Аравийском полуострове [34]. Вполне возможно, что её приход в Танзанию произошёл после 4 тыс. лет назад из Южного Судана с носителями нилотских (нило-сахарских) языков. На это косвенно указывает то, что в Чаде и Камеруне живут представители нижележащей для B-V75 гаплогруппы B-Y12200 (сформировалась 9500 (10600—8500) лет назад, TMRCA 4400 (5200—3700) лет назад), а в Кувейте, Кении и ЦАР носители нижележащей для B-Y12200 гаплогруппы B-BY47577 (сформировалась 3400 (4300—2600) лет назад, TMRCA 3000 (3700—2300) лет назад) [34].
Гаплогруппа E-M2 сформировалась 39200 (42100—36400) лет назад, TMRCA 16400 (18200—14500) лет назад. Представители её дочерней линии E-FT183172 (сформировалась 16400 (18200—14500) лет назад, TMRCA 650 (1100—275) лет назад) живут на Аравийском полуострове, а другой дочерней линии E-Y1705 (сформировалась 16400 (18200—14500) лет назад, TMRCA 14200 (15700—12800) лет назад) в Гамбии и других странах Африки [35]. К сожалению, в данных компании YFull нет жителей Танзании с этой гаплогруппой. Однако, можно предположить, что к сандаве и хадза эта гаплогруппа пришла от бантуязычных лухья, живущих ныне в соседней Кении. В Африку гаплогруппа Е пришла из Азии. Косвенно, об этом можно судить по следующей информации: гаплогруппа E-P116 сформировалась 3600 (4300—2900) лет назад, TMRCA 2900 (4300—1650) лет назад. К базальной линии этой гаплогруппы принадлежит представитель бантуязычного народа лухья в Кении; гаплогруппа E-Z37675 сформировалась 2700 (3400—2100) лет назад, TMRCA 2400 (3800—1400) лет назад. К этой гаплогруппе принадлежат представитель народа лухья из Кении и житель ЦАР; гаплогруппа E-Z1650 сформировалась 2600 (3200—2100) лет назад, TMRCA 2500 (4000—1500) лет назад. Носитель базальной линии этой гаплогруппы живёт в Дубаи, а носители нижележащей линии — в Кении (представители народа лухья) [35].
Гаплогруппа E-M35 сформировалась 34600 (37200—32000) лет назад, TMRCA 23900 (25700—22200) лет назад. Представители одной из её дочерних линий E-L539 (сформировалась 23900 (25700—22200) лет назад, TMRCA 19000 (20800—17300) лет назад) живут в Гамбии и странах Европы, Северной Африки и на Аравийском полуострове (более молодые линии), а другой E-Z827 (сформировалась 23900 (25700—22200) лет назад, TMRCA 23500 (25600—21400) лет назад) в Северной Африке, Чаде, Сомали, Гамбии, странах Европы, на Ближнем Востоке и на Аравийском полуострове [35]. Базальная линия нижележащей для E-M35 гаплогруппы E-Y37091 (сформировалась 1900 (2400—1450) лет назад, TMRCA 1800 (2700—1200) лет назад), определена у жителя Саудовской Аравии и представителя нилотского народа луо, живущего в Кении и Танзании [35].
Ещё одна нижележащая для E-M35 которая может предположительно может быть у сандаве и хадза — гаплогруппа E-Z21175 (сформировалась 4300 (5000—3700) лет назад, TMRCA 4100 (5100—3100) лет назад). Представитель базальной линии её единственной дочерней клады E-CTS7146 (сформировалась 4100 (5100—3100) лет назад, TMRCA 4000 (5100—2900) лет назад), обнаружен в Сомали. Носители одной из дочерних клад E-CTS7146 гаплогруппы E-Y161128 (сформировалась 4000 (5100—2900) лет назад, TMRCA 1500 (2700—750) лет назад) живут в Эфиопии (носитель базальной линии этой гаплогруппы — представитель кушитского народа оромо), Кении (бантуязычный народ лухья), а другой — E-FTA76226 (сформировалась 4000 (5100—2900) лет назад, TMRCA 4000) (5100—2900) лет назад в Омане (араб) и Кении (носители базальной линии), Сомали, Кении, Эфиопии [35].
Таким образом, можно сделать вывод, что у сандаве и хадза в результате контактов с пришельцами, была полностью замещена Y-хромосома (если не появятся новые данные генетических исследований, опровергающие данный вывод).
О том, что в древности митохондриальная гаплогруппа L0 была «спутницей» Y-хромосомной гаплогруппы А00 и эта гаплогруппа была связана с койсанскими народами, косвенно свидетельствуют результаты изучения жителей пустыни Намиб в Анголе, проведённого Oliveira et al. (2018). У них есть данные по народам химба (Himba), кувале (Kuvale), чимба (Tjimba), квепе (Kwepe), Квизи (Kwisi), тва (Twa),!Хун (!Xun) [19, Table S5].
Химба (Himba) — бантуязычный полукочевой народ, занимающийся скотоводством (разведение коз, овец, крупного рогатого скота), ведущий своё происхождение от народа гереро, вместе с которыми мигрировал из Восточной Африки в Намибию, а затем отделился от них [18]. Митохондриальный генофонд химба: L0a1b1 — 0,078, L0a2a1b — 0,013, L0d1b1b — 0,013, L1c1b — 0,091, L1c2b1a — 0,026, L1c3a1b — 0,013, L2c2b1b — 0,013, L2e1 -0,013, L3d3a1a — 0,286, L3e1a2 — 0,234, L3e1a3a -0,026, L3e1e1 — 0,013, L3f`b1a — 0,013, L3f1b4a — 0,169 [19, Table S5]. На гаплогруппу L0 приходится 10,4% генофонда, в то время как на различные субклады L3 — 74,1%.
Кувале (Kuvale) — бантуязычный полукочевой народ, ветвь гереро [27]. По мнению Fehn (2010) язык кувале отличается от языка гереро и должен считаться самостоятельным языком [10]. Митохондриальный генофонд кувале: L0a1b1 — 0,012, L0a1b2 — 0,035, L0a2a1b — 0,012, L0d1a1b — 0,259, L0d1b1b — 0,247, L0d1c3 — 0,024, L1c1b — 0,024, L1c2a1a — 0,035, L1c3b1a — 0,012, L2a1c1 — 0,012, L2c2b1b — 0,024, L2c3 -0,047, L3d3a1 — 0,012, L3d3a1a — 0,012, L3e1a1a — 0,012, L3e2b — 0,024, L3e2b1a — 0,012, L3e4a — 0,012, L3f1b4a — 0,176 [19, Table S5]. На гаплогруппу L0 приходится 58,9%, L3 — 26%.
Если мы обратимся к Y-хромосомному генофонду гереро (Herero), то увидим, что у них нет ни гаплогруппы А, ни гаплогруппы В: E1a — 4,2%, E1b1a — 70,8%, R1b — 12,5%, I — 4,2%, R1a — 4,2% [30]. Это позволяет заключить, что митохондриальная гаплогруппа L0 появилась у гереро после их миграции в Южную Африку.
Чимба (Tjimba) — бантуязычный народ, разговаривающий на языке близком гереро. Соседние народы называют их гереро, потерявшими скот. По-видимому, они являются остатками коренного населения юга Африки, также, как и квади (Kwadi), квизи (Kwisi) и дамара (Damara), которые не похожи на сан (бушмены) и банту гереро [26]. Митохондриальный генофонд чимба состоит только из мтДНК L0 и L3: L0a1b2 — 0,200, L0a2a1b — 0,067, L3d3a1a — 0,133, L3e1a2 — 0,333, L3e2b — 0,133, L3f1b4a — 0,133 [19, Table S5]. На гаплогруппу L0 приходится 26,7%, L3 — 73,2%.
Квепе (Cuepe / Kwepe / Kwadi / Bakoroka / Cuanhoca / Curoca / Koroka / Makoroko / Mucoroca) — практически исчезнувший народ Анголы. Занимались рыбной ловлей и сбором моллюсков. Разговаривают на языке гереро. Последние женщины, знавшие родной язык разговаривали на койсанском языке (Khoi (Click). По-видимому, они остатки юго-западно-африканских охотников-собирателей [8]. Митохондриальный генофонд квепе: L0a1b1 — 0,27, L1c1b — 0,243, L3f1b4a — 0,486 [19, Table S5].
Квизи (Kwisi) — народ, разговаривающий на диалекте языка гереро. Береговые охотники на морскую рыбу и охотники-собиратели Юго-Западной Анголы. По-видимому, являются остатками коренного населения, также как квади (Kwadi), чимба (Tjimba) и дамара (Damara), которые не похожи на койсан (бушменов) и банту [14]. Митохондриальный генофонд квизи: L0a1b1 — 0,25, L0a1b2 — 0,167, L0a2a1b — 0,167, L0d1b1 — 0,042, L1c1b — 0,333, L3f1b4a — 0,042 [19, Table S5]. На гаплогруппу L0 приходится 62,6%.
Тва ((Batwa, Cwa IPA /tʃwa/)) — группа бантуязычных пигмеев. Как и все другие африканские пигмеи, тва, имеют рост в среднем около 1,5 м (4 фута 11 дюймов). Народ смешанного происхождения, вероятно, потомки первоначальных жителей экваториального тропического леса [28]. Митохондриальный генофонд тва: L0a1b1 — 0,167, L0a1b2 — 0,056, L0a2a1b — 0,444, L0d1b1 — 0,056, L1c1b — 0,111, L3f1b4a — 0,167 [19, Table S5]. На гаплогруппу L0 приходится 72,3%.
В нескольких источниках есть данные по Y-хромосомному генофонду пигмеев. Пигмеи мбути (Mbuti): A — 2%, B — 59%, E1b1a — 34%, E2 — 4% [30].
Пигмеи бака (Baka): A — 9,1%, B2b — 63,6%, E* — 12,1%, E1b1a — 12,1%, R1b1* — 3% [6, Fig.1].
Пигмеи бакола (Bakola): B2a — 18,2%, B2b — 18,2%, E* — 4,5%, E1b1a — 54,5%, R1b1* — 4,5% [6, Fig.1].
Согласно этим данным, основная Y-хромосомная гаплогруппа пигмеев и судя всему их первоначальная, родная — В.
!Хун (!Xun) — бушменское племя, в основном проживающее на западе пустыни Калахари, Овамболенде и Ботсване. Митохондриальный генофонд! Xun: L0d1b1a — 0,026, L0d1b2a — 0,026, L0d1b2b — 0,026, L0d1c1a — 0,256, L0d1c1a — 0,103, L0d2a1a — 0,359, L0ka1 — 0,051, L0ka1a — 0,077, L0k1a1b — 0,051, L3d3a1 — 0,026 [19, Table S5]. На митохондриальную гаплогруппу L0 приходится 97,5%. При этом, наибольшую частоту имеют субклады гаплогруппы L0d — 79,6%
По койсанским народам также есть данные по Y-хромосоному генофонду. Койсаны Южной Африки (Khwe,!Kung, mixed): A — 33,3%, B — 12,4%, E1b1a — 35,7%, E1b1b — 14,7%, E2 — 3,9% [25].
Койсаны ǃKung: A — 36%, B — 8%, E1b1a — 39%, E1b1b — 11%, E2 — 6% [7].
Койсаны (!Kung/Sekele, Tsumkwe San, Dama, Nama): A-P3* — 14,4%, A-P28 — 11,1%, A-M51 — 22,2%, B-M182* -1,1%, B-P6 — 8,9%, B-P7 — 4,4%, E-SRY₄₀₅₄* — 1,1%, E-M75* — 1,1%, EM54 — 1,1%, E-P1* — 14,4%, E-M191 — 10,0%, E1b1b-M35* — 6,7%, J12f2 -1,1%, R-M269 — 2,2% [30]. Отдельно по племенам:!Kung/Sekele: A-M6 (xA-P28) — 16%, A2b-P28 — 9%, A1b1b2a (M51; formerly A3b1) — 22%; Tsumkwe San: A-M6 (xA-P28) — 28%, A2b-P28 — 17%, A1b1b2a (M51) — 21%; Dama: A2b-P28 — 9%, A1b1b2a (M51) — 6%; Nama: A2b-P28 — 9%, A1b1b2a (M51) — 55% % [30, Appendix A].
A-M6 — это A-Y25057 (сформировалась 25300 (27800—22800) лет назад, TMRCA 12100 (14000—10200) лет назад) [33]. Согласно данным компании YFull обе её дочерние клады A-P28 и A-L963 обнаружены на юге Африки (Ангола, Намибия) [33].
A-M51 сформировалась 53200 (56800—49700) лет назад, TMRCA 16300 (18600—14100) лет назад. Носитель её базальной линии обнаружен в Анголе, а представители её единственной дочерней клады A-M9782 (сформировалась 16300 (18600—14100) лет назад, TMRCA 14900 (17000—12900) лет назад), живут в ЮАР, Анголе и Намибии. Представители сестринской для А-М51 гаплогруппы A-YP4735 живут в Восточной Африке и в подаляющем большинстве на Аравийском полуострове [33].
Таким образом, приведённые данные, скорее всего, свидетельствуют в пользу замещения у койсанских народов Y-хромосомной гаплогруппы А00 гаплогруппой А1.
V. Гаплогруппа А0-Т и создатели археологических индустрий/культур, которые могут претендовать на роль её носителей
Гаплогруппа А0-Т сформировалась 235900 (243700—228300) лет назад TMRCA 161300 (169900—152900) лет назад. Имеет две дочерние клады: A0 (сформировалась 161300 (169900—152900) лет назад, TMRCA — не рассчитан), по-видимому, только африканскую и А1 (сформировалась 161300 (169900—152900) лет назад, TMRCA 133400 (139000—127800) лет назад), одна дочерних субклад которой (A1b1), в основном, африканская, а другая (ВТ) — широко распространена в мире [72].
На роль популяции, которую можно ассоциировать с носителями гаплогруппы А0-Т могут претендовать создатели индустрии Табун D из Леванта, люди из формации Гадемотта/Кулкулетти из Восточно-Африканской рифтовой долины в Эфиопии (Main Ethiopian Rift Valley) и эфиопских стоянок Омо Кибиш KHS (Omo Kibish KHS), Омо Кибиш AHS (Omo Kibish AHS), Херто (Herto), выделенных Zaidner et al. (2021) в группу «Индустрия MSA: Центростремительное леваллуа, связанное с редкими ручными рубилами и/или бифасами» (MSA industry: Centripetal Levallois associated with rare handaxes and/or bifaces) [73, Table S1].
5.1. Табун D, Мислия 1 и гаплогруппа А0-Т
Можно предположить, что развитие людей современного анатомического типа — носителей Y-хромосомных гаплогрупп A0-T, А0 и А1, происходило в рамках левантийского мустье фации Табун D. Данное предположение основано на трёх предположениях:
1) совпадение периодов существования индустрии Табун D и Y-хромосомной гаплогруппы А0-Т;
2) в пещере Мислия, индустрия которой относится к Табун D, были найдены самые ранние в Леванте антропологические окаменелости, относящиеся, по заключению Hershkovitz et al. (2018), к людям современного типа [37];
3) наличие археологических культур инициального верхнего палеолита, по предположению исследователей, являющихся потомками Табун D;
4) благоприятные климатические условия, обусловившие возможность миграций из Африки в Азию и обратно через Синайский полуостров, во временном интервале между 246—200 тыс. лет назад [24].
5.1.1. Общая характеристика индустрии Табун D
К фации Табун D археологи относят индустрии стратиграфического подразделения D пещеры Табун в Кармеле, а также местонахождений Рош-эйн-Мор (Rosh Ein Mor) и Нахаль Акев (Nahal Aqev) в горах Негев, Хайоним (слои Е (низ) и F) в Западной Галилее, Абу-Сиф (Abu Sif) и Сахба (Sahba) в Иудейской пустыне, Джерф Айла (Jerf Ajla) и Дуара IV (Douara Layer IV) в Восточной Сирии, Айн Дифла (Ain Difla) в западно-центральной части Иордании, Хуммал, Мислия и др. [8, с. 277, 287; 36, p.14].
Можно отметить, что информация о технологии изготовления каменных орудий на стоянках Табун D противоречива. Так, например, согласно Монигал (2001) в Табун D применялись однонаправленное, двунаправленное и центростремительное расщепление [16, с. 17].
Zaidner et al. (2021) утверждают, что для индустрий Табун D характерно сочетание леваллуазской и пластинчатой технологий для изготовления удлинённых леваллуазских наконечников, а также треугольных отщепов и пластинок, различных ретушированных наконечников. Однако несмотря на присутствие технологии леваллуа на памятниках Табун D, на них практически отсутствует центростремительное леваллуа. В пещерах Хайоним и Мислия частота встречаемости центростремительных отщепов Леваллуа колеблется от 1,8 до 3,4% [73]. По мнению Zaidner et al. (2021), небольшое количество таких отщепов может быть случайным результатом любой последовательности расщепления и не обязательно результатом центростремительного метода Леваллуа, их присутствие в небольшом количестве на участках EMP не может рассматриваться как однозначный показатель использования центростремительного метода Леваллуа на этих стоянках [73].
Henry (2003) пишет, что в хуммалиене применялась пластинчатая система, но отсутствует метод леваллуа [36, p.14]. Хуммал расположен возле д. Эль-Ковм (El-Kowm) в центральной Сирии, между бассейном р. Евфрат и пустынной степью, простирающейся от Пальмиры в Дейр-эз-Зор (Deir ez-Zor). Индустрия среднепалеолитических слоёв 6а-6в и 7 этого памятника отнесена к хуммалиену [70]. Wojtczak (2015) пишет, что в хуммалиане представлены нуклеусы на отщепах, выполненные в технике Нар Ибрагим или без неё. С индустрией хуммалиен демонстрируют наибольшее сходство индустрии из слоёв Хайонима F и E, недатированных слоёв Абу Сиф B и C, пещер Мислия и Эмануэль [69, p.34].
Hershkovitz et al (2018) отмечают, что для индустрии Табун D характерна развитая однополярная леваллуазская технология, основанная на производстве удлинённых заготовок (пластин и леваллуазских острий). На ряде стоянок также использовались другие стратегии редукции нуклеусов. Например, на стоянках Рош-Эйн-Мор (Израиль) (около 200 тыс. лет назад), в пещере Мислия на склоне горы Кармель, в 12 километрах к югу от Хайфы (250—150 тыс. лет назад) применялась леваллуа-пластинчатая традиция. Технологические концепции, подобные применяемой в Мислии, были задокументированы на современных и более ранних стоянках среднего палеолита в Африке, — в Магрибе (Джебель-Ирхуд), Восточной Африке (формации Гадемотта (Gademotta) и Кулкулетти (Kulkuletti), Эфиопия, формация Каптурин (Kapthurin), Кения)) и в Южной Африке (Кату-Пан (Kathu Pan)) [37].
Выше было показано, что индивид Джебель-Ирхуд-1 из магрибского местонахождения Джебель Ирхуд, скорее всего, Homo heidelbergensis [2, с. 105]. Соответственно, создатели индустрий, перечисленных Hershkovitz et al. (2018) могут принадлежать к этому древнему виду человека.
5.1.2. Совпадение периодов существования индустрии Табун D и Y-хромосомной гаплогруппы А0-Т
Гаплогруппа A0-T сформировалась 235900 (243700—228300) лет назад, TMRCA 161300 (169900—152900) лет назад. Её дочерние клады: A0 — сформировалась 161300 (169900—152900) лет назад; A1 — сформировалась 161300 (169900—152900) лет назад, TMRCA 133400 (139000—127800) лет назад [72].
Датировки Табун D согласно разным авторам: ЭПР-даты индустрии типа Табун D — 230—120 тыс. лет назад, термолюминесцентные — 280—230 тыс. лет назад [5, с. 44]; леваллуа-мустьерская индустрия слоя D пещеры Табун –263–244 тыс. лет назад [6, с. 20].
Табун D методами электрон-спин-резонанса и урановых серий датирован 133—203 тыс. лет, термолюминисценции — 263—270 тыс. лет, с границей со слоем С около 244 тыс. лет [10, с. 47]; пещера Мислия — минимальная ОСЛ дата 130±33 тыс. лет назад, стоянка Айн Дифла — ТЛ дата 105±15 тыс. лет назад и ЭПР даты от 102±13 до 162±18 тыс. лет назад, индустрия нижних слоев стоянки Бокер Тахтит — не более 50 тыс. лет (радиоуглеродные даты) [3, с. 60—61].
Valladas et al. (2013) среднепалеолитическую индустрию в пещере Мислия датируют периодом 250–140 тыс. лет назад [65].
Мустьерские слои 5a-5g в Хуммале имеют термолюминисцентные даты (TL) ~35-~100 тыс. лет назад; хуммалиен (Hummalian) (слои 6—7) датируется ~200 лет назад (TL) [70, Table 1, p.181]. Нужно отметить, что возраст хуммалиена термолюминесцентным методом (TL) был определён в 104±9 тыс. лет. Однако, по мнению Wojtczak (2015), это заниженная дата и предполагаемый возраст хумаллиена ~200 тыс. лет, который соотносится с возрастами аналогичных пластинчатых индустрий раннего среднего палеолита (EMP) — слои E (верх) и F (низ) Хайонима со средними TL-датами 210±28 тыс. лет назад и 221±21 тыс. лет назад соответственно, полученными при нагреве кремния; пачки Табун IX (тип Табун D), где тот же метод даёт 256±26 тыс. лет назад; пещеры Мислия со средним возрастом варьирующим от 212±27 до 166±23 тыс. лет назад; пещеры Эмануэль (Emanuel Cave), датированной уран-ториевым (U/Th) методом 191 тыс. лет назад; Айн-Дифла (Ain Difla) датируемой примерно между 90 и 180 тыс. лет назад и Рош-Эйн-Мор (Rosh Ein Mor), датируемым 200 тыс. лет назад [69].
Таким образом, можно констатировать, что датировки фации Табун D неплохо соотносятся с возрастом Y-хромосомной гаплогруппы A0-T.
5.1.3. Мислия 1 — человек современного анатомического типа?
Пещера Мислия (пещера Броцена) расположена на западном склоне горы Кармел, в 12 км к югу от г. Хайфы и примерно в 7 км к северу от пещер Табун, Джамаль, Эль-Вад и Схул. На нижней террасе пещеры были обнаружены ашело-ябрудьенский и среднепалеолитический культуросодержащие слои, на верхней — раннего среднего палеолита.
Ашело-ябрудийская индустрия из пещеры Мислия, датируемая концом МИС 8 — началом МИС 7 (MIS 7 — это период, охватывающий 246–186 тыс. лет назад [67]), отражает три технологии расщепления: бифасиальную; скалывания тонких отщепов с подготовленных нуклеусов; снятия крупных массивных часто первичных сколов с неподготовленных нуклеусов, служивших заготовками рубил, скрёбел кина и полукина. В целом для этой индустрии Мислии характерно преобладание отщепов в числе сколов, присутствие редких пластин, отсутствие леваллуазских изделий, а также наличие удлинённых заготовок неправильной формы, без признаков преднамеренного пластинчатого расщепления [14, с. 46].
При раскопках пещеры Мислия были найдены самые ранние в Леванте антропологические окаменелости, относящиеся, по заключению Hershkovitz et al. (2018), к людям современного типа: часть верхней челюсти (с неповрежденными I²–M³), четыре отдельных зуба, фаланга и коленная чашечка. Обнаружены также остатки очагов и свидетельства постоянного использования обитателями пещеры огня [37].
Первичное расщепление в пещере Мислия по большей части представлено леваллуазской системой получения заготовок в виде остроконечников, среди которых преобладали удлинённые формы (коротких остроконечников с широким основанием обнаружено немного), а также укороченных отщепов. Индустрия на стоянке Мислия отличается частым использованием пластин в качестве заготовок для изготовления орудий. Пластины скалывались с призматических и поворотных пластинчатых нуклеусов. В составе каменного инвентаря отщепы численно преобладали над пластинчатыми заготовками, однако для производства орудий пластинчатые заготовки применялись гораздо чаще. Из леваллуазских нуклеусов наиболее многочисленны на стоянке треугольные и подтреугольные в плане. С них производилось скалывание однонаправленных конвергентных заготовок. Униполярные нуклеусы имели хорошо подготовленную, часто фасетированную ударную площадку. Аналогичная система оформления остроконечных леваллуазских заготовок выявлена и на других стоянках раннего среднего палеолита в Леванте, например, в Табун D [8].
Ближайшие аналогии среднепалеолитической индустрии в пещере Мислия можно проследить в слое Е пещеры Хайоним и в индустрии стоянки Хуммал (бассейн Эль-Коум в Сирии).
Факт принадлежности Мислия-1 к современным людям, установлен по части верхней челюсти (половина левой части верхней челюсти со всеми зубами, от первого резца (от него остался только корень) до третьего коренного). Компьютерная томография и сравнение её строения с образцами древних и современных челюстей, показали, что по форме зубной дуги, строению скулового отростка, ориентации дна носовой полости и другим особенностям, челюсть Мислии-1 приближается к современным людям, а также древним сапиенсам типа Джебел Ирхуд и Херто. Анализ строения зубов показал, что у Мислия-I резцы вполне сапиентные, клыки не имеют особенностей зубов эректусов и неандертальцев и похожи на зубы Схул и Кафзех; это же относится и к предкоренным зубам. Двухмерный морфометрический анализ коронки первого коренного зуба расположил этот зуб рядом с зубом Джебел Ирхуд. Зубы Мислия-1 по размерам находятся в верхних границах изменчивости современных людей [37].
Челюсть датируется возрастом 174±20 тыс. лет назад (календарных) (при стандартном отклонении 2σ), полученным при совместном применении методов урановых рядов (Uranium-series, U-series) и электронно-парамагнитного резонанса (ЭПР), с орудиями, характерными для фации Табун D, в пещере Мислия (Misliya Cave) [37].
Поскольку зуб Мислии близок к зубу Джебель-Ирхуда 1, который, скорее всего, Homo heidelbergensis, то и Мислия, может принадлежать к этому древнему виду человека. Кроме того, ниже будет рассмотрена информация о человеческих останках из левантийской пещеры Нешер Рамла. Индустрия пещеры относится к мустье Табун С и центростремительному леваллуа, а её обитатели к отдельному виду Nesher Ramla Homo [74], который сформировался на базе более древних Homo heidelbergensis. Поскольку Мислия 1 отнесли к анатомически современным людям, изучив челюсть, то может оказаться, что она относится к тому же архаичному виду, что и люди из пещер Нешер Рамла, Кесем и др.
Нужно также учитывать, что Sharp, Paces (2018) утверждают, что у Hershkovitz et al. (2018) нет достоверных U-series дат старше ~70 тыс. лет назад и останки Мислия имеют минимальный возраст не старше ~60—70 тыс. лет [58]. Поэтому Мислия может оказаться человеком современного анатомического типа, жившего 60—70 тыс. лет назад.
5.1.4. Происхождение среднепалеолитической индустрии Табун D
Valladas et al. (2013) заметив ряд различий индустрий финального этапа нижнего палеолита (ашело-ябрудийен) и раннего этапа среднего палеолита в пещере Мислия (Табун D), пришли к выводу, что около 250 тыс. лет назад (конец MIS 8) в Левант пришла новая популяция [65]. По их мнению, она могла быть связана либо с неандертальцами из Европы, либо с людьми современного анатомического типа из Африки [65, p.592].
Деревянко (2016) отрицает возможность прихода обитателей пещеры Мислия из Африки и Европы. Обосновывает он это положение тем, что позднеашельская пластинчатая индустрия формации Каптурин в Восточной Африке, датируемая около 280 тыс. лет назад, не имеет никаких аналогий с ашело-ябрудийской индустрией Леванта, а средний каменный век Африки также отличается от среднего палеолита Леванта. Поэтому, нельзя говорить о проникновении из Африки в Левант людей современного анатомического типа около 250 тыс. лет назад. В Европе также неизвестны мустьерские культуры, которые могли бы послужить основой формирования комплексов Табун D. Он утверждает, что гораздо «больше свидетельств того, что истоком среднепалеолитического технокомплекса Леванта являлась ашело-ябрудийская индустрия» [7, с. 20]. В Леванте, по мнению Деревянко (2016), происходила сапиентация популяций, заселивших эту территорию в среднем и первой половине верхнего плейстоцена, «в результате чего здесь сформировались два таксона, представленные людьми современного анатомического вида и палестинскими, или переднеазиатскими, неандертальцами» [7, с. 20—21]
Таким образом, если прав Деревянко (2016), создатели индустрии Табун D — гейдельбергский человек (Homo heidelbergensis) или Homo bodoensis, если пользоваться терминологией Roksandic et al. (2022) [53].
5.1.5. Археологические культуры/индустрии — возможные потомки Табун D
Деревянко (2016) считает, что «Индустрия среднего палеолита Леванта типа Табун С истоками была связана с технокомплексом раннего периода среднего палеолита типа Табун D и в своём развитии не претерпела коренных изменений, что должно было произойти при аккультурации с приходом популяции людей современного анатомического вида и тем более при замещении автохтонного населения пришлым» [7, с. 20].
С ранним левантийским мустье типа Табун D ассоциируется появление платинчатой микроиндустрии [49] и предполагается связь населения Табун D с распространением леваллуа-пластинчатых индустрий в Евразии [17].
Кривошапкин (2012) полагает, что из ближневосточного мустье типа Табун D происходит технология получения пластин с плоскостных и объёмных нуклеусов, характерная для Оби-Рахмата (долина реки Пальтау, Бостанлыкский район Ташкентской области) в Узбекистане [15, с.27—28].
Группа российских археологов под руководством А.И.Кривошапкина проведя статистический анализ артефактов-маркеров в комплексах стоянок Средней Азии — Кульбулак (слой 23, 2010 г.), Оби-Рахмат (слои 4—14) и Леванта — Хайоним F, Эль Ковм, установила, что обирахматские комплексы показывают значительную долю сходства с ранним левантийским мустье.
Атрибутивный анализ слоёв 15—19 грота Оби-Рахмат показал их технико-типологическую преемственность между собой. Близкие характеристики продемонстрировал комплекс слоя 24 стоянки Кульбулак. Была доказана общность орудийных коллекций комплексов грота Оби-Рахмат (слои 2—19), стоянки Хонако-3, ПК2 и индустрии стоянки Кульбулак, слой 23 (2010 г. раскопок).
Для стоянки лёссового палеолита Хонако 3 получены абсолютные даты. Верхние 5 метров разреза сформировались в период между 23—41 тыс. лет назад. Нижняя часть дат характеризует возраст ПК2 — 180—220 тыс. лет назад. Даты для второй палеопочвы с обирахматским комплексом показывают самое раннее проявление пластинчатой среднепалеолитической традиции в регионе.
Исходя из того, что пластинчатые обирахматские индустрии, существовали, по предварительным данным, в интервале 120—45 тыс. лет назад, исследователи выдвинули гипотезу о непрерывном, растянутом географически, миграционном процессе, в результате которого популяция ранних Homo sapiens sapiens с территории Ближнего Востока принесла в западную часть Центральной Азии среднепалеолитическую пластинчатую индустрию [11].
Pavlenok et al. (2024) отмечают сходство подготовки нуклеусов для производства небольших пластин в археологических комплексах Леванта ~240–150 тыс. лет назад (пещера Мислия (Misliya Cave); пещера Хайоним, слой F (layer F of the Hayonim Cave) и др.) и Средней Азии (Кульбулак слой 23; индустрия Оби Рахмата, датируемая 80 тыс. лет назад). По их мнению, эта технологическая традиция получила дальнейшее развитие в верхнем палеолите Средней Азии (MIS 2 — вторая половина MIS 3) [52].
По мнению Рыбина (2020), появление сходных типичных пластинчатых индустрий начального верхнего палеолита в Леванте и на Алтае является синхронным и, возможно, связано с «развитием на общей основе предковых леваллуа-пластинчатых индустрий Леванта типа раннелевантийского мустье представленных в комплексах Мислия и Рош-эйн-Мор» [18, с.498].
По мнению Marks (1983), комплексы раннего левантийского мустье существовали вплоть до перехода к верхнему палеолиту — из мустье типа Табун D, продолжавшего развиваться в южных районах Леванта (пустыня Негев, Израиль) до 60—50 тыс. лет назад, происходит эмиран [46], который в настоящее время считается источником инициального верхнего палеолита.
Barzilai et al. (2022) выводят эмиран из пустыни Негев, где, по их мнению, произошло смешение носителей левантийского (Табун D) и нубийского среднего палеолита [22].
Коен и Степанчук (2001) прослеживают признаки технокомплекса Табун D, в кременецкой индустрии нижних слоёв стоянки Кулычивка в Западной Волыни, датируемых около 31 тыс. лет назад [13, с.90—91]. При этом они отмечают, что нижний слой Кулычивки гетерогенен и представлен орудиями, типичными для технологически сходных индустрий (Богунисьен, Темната слой VI, Эмиран, а также с определёнными оговорками леваллуа-мустье типа Табун D), а также ориньякским типологическим компонентом [13, с.91].
Демиденко (2020) относит к раннему левантийскому мустье типа Табун D, индустрии расположенных на Кавказе и в Закавказье пещер Кударо I, III и Цона (Южная Осетия); пещеры Джручула в Имеретии (Западная Грузия); пещеры Ховк I и стоянки открытого типа Баграташен I в Северной Армении [4, с.131].
5.2. Гадемотта/Кулкулетти (Gademotta/Kulkuletti)
Гадемотта и Кулкулетти (Gademotta and Kulkuletti) — два, расположенных по соседству местонахождения под открытым небом, относящиеся к раннему среднему палеолиту, находящиеся на склонах над западным берегом озера Зивай (Ziway) в геологической формации Гадемотта, в Центральной рифтовой долине (Central Rift Valley), в Эфиопии (рис. 19).
Рис. 19. Расположение Гадемотта и Кулкулетти [32]
Morgan, Renne (2008) утверждают, что в отличие от таких памятников, как Каптурин, Омо Кибиш, Херто, в которых комплексы раннего MSA содержат также ашельские артефакты, ассамбляжи Гадемотта состоят из орудий, которые технологически и типологически относятся исключительно к MSA, датируемому 276±4 тыс. лет назад (самый старый участок формации Гадемотта был датирован в 2008 году аргон-аргоновым методом (⁴⁰Ar/³ꝰAr) — 276 тыс. лет назад. Эта дата — наиболее старая для памятников MSA со сходной технологией) [50]. Ассамбляжи MSA Гадемотты, датированные ~276 тыс. лет назад, содержат значительный пластинчатый компонент [40, p.194].
Получена также дата 183 тыс. лет назад для разреза, которая по мнению Morgan, Renne (2008), указывает на технологический застой, наблюдаемый по всему разрезу, и на то, что аналогичная технология использовалась здесь на протяжении ~93 тыс. лет [50].
Sahle et al. (2013) установили, что заостренные обсидиановые артефакты, датируемые> 279 тыс. лет назад (дата установлена аргон-аргоновым методом; ⁴⁰Ar/³ꝰAr), обнаруженные на стоянках формации Гадемотта (Fm), представляют собой наконечники составного метательного оружия — копья. Вместе с тем, исследователи считают, что в этот период (> 279 тыс. лет назад) Homo sapiens не существовал, т.е. это оружие создал не анатомически современный человек [54].
В формации Гадемотта, также, как и в формации Каптурин, задокументированы самые ранние свидетельства центростремительной леваллуазской технологии в Северо-Восточной и Восточной Африке. Здесь обнаружены центростремительные отщепы леваллуа, центростремительные преференциальные и рекуррентные нуклеусы леваллуа, отщепы débordant, а также псевдолеваллуазкие отщепы и острия [73]. Забегая вперёд можно упомянуть результаты исследования Zaidner et al. (2021) которые установили, что самые ранние появления Homo sapiens в Юго-Западной Азии (~180 тыс. лет назад) связаны со среднепалеолитическими индустриями с леваллуазской технологией. На стадии MIS 5 все окаменелости Homo sapiens в Западной Азии были связаны с центростремительным методом Леваллуа. Поэтому эту технологию иногда используют как маркер распространения Homo sapiens в Западную Азию во время MIS 5 [73].
Douze, Delagnes (2016) изучив технологию изготовления конвергентных каменных орудий формации Гадемотта/Кулкулетти за период от 280 тыс. лет назад до около 100 тыс. лет назад, пришли к мнению, отличающемуся от точки зрения Morgan, Renne (2008), которые утверждают о технологическом застое в формации Гадемотта/Кулкулетти в период от 276±4 до 183 тыс. лет назад [50]. Они выделили важные диахронические изменения в процессе изготовления конвергентных инструментов:
1) разработка специфических леваллуазских методов производства острий (points) (классическое производство острий леваллуа и редукция нубийских нуклеусов типа 1);
2) переход от одно- двуфациального (uni-bifacial) инвазивного придания формы конвергентным инструментам к локализованной лёгкой ретуши в заранее определённых точках [33]. По мнению Douze, Delagnes (2016), эти технологические изменения в производстве конвергентных инструментов свидетельствуют о постепенном появлении нового набора технологических моделей поведения, которые можно считать специфичными для MSA [33].
Для характеристики создателей индустрий африканского среднего палеолита важны результаты работы McCall (2015), который исследовав каменные технологии MSA Гадемотта/Кулкулетти и Омо Кибиш утверждает, что характерные черты каменных каменных комплексов MSA связаны с системами расселения. Гоминины MSA при поисках пищи использовали стратегию мобильности, совершая частые переходы и занимая жилые участки на короткие периоды времени, чем отличаются от современных людей-собирателей. Эта модель имела последствия для перехода от ашельского периода к MSA [47].
Hershkovitz et al (2018) отмечают сходство с индустрией формации Гадемотта/Кулкулетти индустрии Табун D (методами электрон-спин-резонанса и урановых серий датирована 133—203 тыс. лет, термолюминисценции — 263—270 тыс. лет, с границей со слоем С около 244 тыс. лет [10, с. 47]) из Леванта [37]. При этом, они, по-видимому, имеют ввиду индустрию Гадемотта/Кулкулетти, датируемую периодом 276—183 тыс. лет назад. Если это так, то вероятно, что создатели индустрии Табун D могут иметь общее происхождение с популяцией из формации Гадемотта/Кулкулетти.
Если правы Hershkovitz et al. (2018) в том, что индустрия Джебель Ирхуда, созданная скорее всего Homo heidelbergensis [2, с. 105], сходна с индустрией Табун D из Леванта, а индустрия Табун D сходна с каменными орудиями формации Гадемотта/Кулкулетти, датируемыми 276—183 тыс. лет назад [37], то, возможно, индустрии и Табун D и формации Гадемотта/Кулкулетти этого периода — творение Homo heidelbergensis. Однако нельзя забывать, что африканский гейдельбергский человек считается предком современного человека (Homo sapiens sapiens) [1; 12, с. 103] (по версии Roksandic et al. (2022) — гейдельбергский человек Африки — это Homo bodoensis [53]).
Поэтому, исходя из дат, более вероятен вариант, что обитатели формации Гадемотта/Кулкулетти (и других стоянок в данном регионе) — люди современного анатомического типа, точнее — прямые предки большей части современного человечества.
Исходя из вышеизложенного (и материалов по мтДНК) можно сделать выводы:
1) 276±4 тыс. лет назад в формацию Гадемотта/Кулкулетти откуда-то извне пришла группа людей, которая, как установил McCall (2015) была очень подвижной, совершая частые переходы в поисках пищи и занимая жилые участки на короткие периоды времени [47].
Если опираться на результаты исследования Schlebusch et al. (2017) согласно которому Homo sapiens появились на Земле ранее 300 тыс. лет назад (самое раннее расхождение между человеческими популяциями по результатам их исследования могло произойти от 350 до 260 тыс. лет назад) [56], то это могли быть люди современного анатомического типа, носители или Y-ДНК А0-Т или А00, которые имеют возраст 235900 (243700—228300) лет назад [72]. Данный вывод подтверждается археологическими свидедельствами — каменная индустрия в Гадемотта/Кулкулетти представлена орудиями, которые технологически и типологически относятся исключительно к MSA, датируемому 276±4 тыс. лет назад [50].
Рис. 20. Карта Восточной Африки, показывающая расположение объектов MSA: 1. Abdur; 2. Aduma; 3. Aladi Springs; 4. Bodo; 5. Cartwright’s Site; 6. Deighton’s Cliff; 7. Enkapune ya Muto; 8. Gademotta/Kulkuletti; 9. Gorgora rock shelter; 10. Herto Member; Douri Fm.; 11. K’one; 12. Kapedo Tuffs; 13. Kapthurin Fm.; 14. Kibish Fm.; 15. Kisese II rock shelter; 16. Koobi Fora; 17. Lake Eyasi Beds; 18. Loyangalani; 19. Lukenya Hill; 20. Midhishi 2; 21. Moche Borago; 22. Mtongwe; 23. Muguruk; 24. Mumba rock shelter; 25. Nasera rock shelter; 26. Njoro River Cave; 27. Olorgesailie; 28. Porc Epic; 29. Prolonged Drift; 30. Prospect Farm; 31. Rambogo rock shelter; 32. Rusinga Island; 33. Songhor; 34. Upper Ngaloba Beds, Laetoli [63, Fig. 1]
2). На рис. 20 показано, что местонахождения MSA образуют две группы памятников. Одна из них располагается вблизи озера Туркана и к северу от него, а вторая группируется вблизи озера Виктория и на восток от него к побережью Индийского океана. Первая группа памятников включает местонахождения формаций Кибиш, Гадемотта/Кулкулетти, формации Боури (Херто), памятники Адума, Порк Эпик и др.; вторая группа — памятники формации Каптурин, Мугурук и др. Исходя из того, что в настоящее время носители Y-хромосомной гаплогруппы А0-Т занимают территорию первой группы памятников, а также Северную Африку, Евразию и Америку, логично предположить, что стоянки первой группы связаны с носителями Y-хромосомной гаплогруппы А0-Т и её нижележащих линий. Вторую группу стоянок в таком случае можно отождествить с носителями Y-хромосомной гаплогруппы А00.
Бесплатный фрагмент закончился.
Купите книгу, чтобы продолжить чтение.